
古生物简讯(2023第一季度)
大家好,我是黑莓,我们好久不见了。
转眼间,2023年已经过去了四分之一,过去的3个月里,以《史前星球》和《恐龙超世界》为代表的多部纪录片纷纷放出了新作或是续作的预告片,并将在不久之后与大家见面;同时,各个领域也陆陆续续的公布了各种各样的新研究,在这里,我将挑选15个比较有代表性的研究进行简单的讲解复盘。
那么废话不多说,我们现在开始。

鸟臀目演化的深层差异
1月4日,期刊《Current Biology》刊登了英国科学家对鸟臀目演化深层差异的研究,在研究中,他们选取了鸟臀目多个分支中的基干类群,包括畸齿龙(Heterodontosaurus)、莱索托龙(Lesothosaurus)、腿龙(Scelidosaurus)、棱齿龙(Hypsilophodon)以及鹦鹉嘴龙(Psittacosaurus),并对它们的头骨进行有限元分析,重建颞区肌肉以探讨它们的形态功能学。
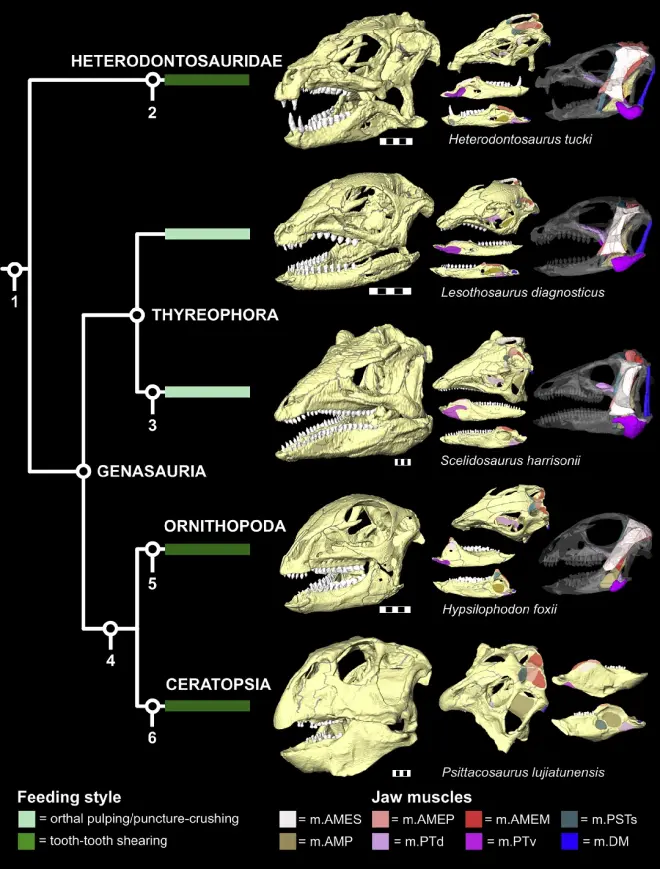
研究显示:它们不同位置的绝对咬合力都拥有很大的差异,这其中莱索托龙的绝对咬力和相对咬力都较低,而鹦鹉嘴龙的相对咬合力和绝对咬合力都很高。至于其它的类群:棱齿龙和畸齿龙都和莱索托龙一样拥有较低的绝对咬合力,但棱齿龙的相对咬合力较高;腿龙则是相对咬合力和绝对咬合力都处在中等较高的水平上。
不同类群可以通过明显不同的方式来达到相同的咬合效率。举例来说:畸齿龙颌骨收肌的绝对面积较小,但它相比于莱索托龙和棱齿龙拥有更大的相对颌骨收肌面积,畸齿龙也以此提升了自身的咬合效率;相对的,腿龙依靠较大的体型达到了较高的绝对颌骨收肌面积,但其相对面积更小,所以二者的咬合效率反而没有明显差距。而鹦鹉嘴龙则是一个完全的异类:无论是绝对还是相对面积都相当高,因此无论是咬合力还是咬合效率都非常的高。

应力分析显示:鹦鹉嘴龙的头骨和下颌在咬合的过程中受到的应力是很小的,畸齿龙的头骨则受到了更大的应力,莱索托龙和棱齿龙拥有类似的应力分布模式以及强度,而腿龙在利用下颌末端咬合时应力下降最为明显。
通过 一系列的研究,科学家证明这些基干类群的趋同是十分有限的,它们通过不同方式适应饮食,并获得了类似的功能:装甲类通过增加头骨绝对尺寸、畸齿龙类通过扩张颌骨收肌尺寸、鸟脚类提高颌骨使用效率、角龙类则将这些特征集合起来。鸟臀目各分支基干类群的不同适应手段反映了它们对环境的解决方案多样化,这将有助于它们的存活和发展。同时,此次的结果也与之前的一些研究相对应,例如此前的研究表明莱索托龙可能更加偏向杂食,而这次的研究结果也在一定程度上证明了它并不像其它以坚硬植物为食的恐龙拥有发达的咬合力。

古老的吸食性全头类
与现在形态学上相对保守的底栖硬食性银鲛不同,古生代的全头亚纲(Holocephali)拥有惊人的多样性,它们当中的不少类群都拥有十分奇特的解剖结构,1月17号,期刊《Proceedings of the National Academy of Sciences of the United States of America》刊登了法国的科学家对生活在宾夕法尼亚世美国的袋枕鲛(Iniopera)的研究。袋枕鲛属于枕鳍鲛目(Iniopterygiformes),它曾经被认为属于硬食性,但它缺少类似银鲛适应于硬食的结构,例如齿板以及朝向前下方的嘴巴。得益于保存完好的标本,科学家可以对其骨骼和肌肉进行较准确的3D重建,并借此重新研究其习性。
重建的结果显示:袋枕鲛的下颌收肌直接插入下颌骨,这更类似于鲨类而非银鲛;它的颅顶肌肉附着量要少于银鲛,但枕骨大孔两侧发达的窝表明它拥有发达的肌肉附着。袋枕鲛的胸鳍异常靠向背侧,脑颅上发达的后窝以及基鳃骨和肩胛乌喙骨上较大的肌肉附着点表明它头骨和肩带的肌肉连接十分发达;它的舌骨相比于叶吻银鲛也更加粗壮,喙舌肌发达,表明舌骨在袋枕鲛进食的过程中会发挥十分重要的作用。

进一步研究显示:袋枕鲛的口裂最大可以达到56.5°-63.5°,并在28°-33.5°之间为最佳。由于下颌收肌的位置过于靠后,无法发挥出更多的机械优势,它的咬合力很低,末端最大也只有3.94N,基于这些再结合袋枕鲛靠前的吻部以及缺乏唇侧软骨的嘴唇,它大概率并不是一类硬食性物种。而相对地,它拥有发达的舌骨和基鳃骨、可扩张的咽部以及发达的颅骨肩带肌肉群,这些特征更多的与高效吸食性有关。

大量的证据都证明袋枕鲛很可能是一种吸食性的全头类,但相比于其它的鱼类,它的吸食模式似乎更接近今天的四足动物(例如玛塔龟),拥有愈合的脑颅和上颚以及较短的咽部利用舌骨和肩带扩张咽部进行吸食,而板鳃类使用唇软骨收缩口部,并通过颌骨和舌骨扩大口腔容积。
袋枕鲛可能占据着类似于现在新鳍鱼类的生态位,表明全头亚纲在辐鳍鱼类大规模多样化之前曾经占据着比我们想象中更加广阔的生态位。

奇特的滤食性翼龙
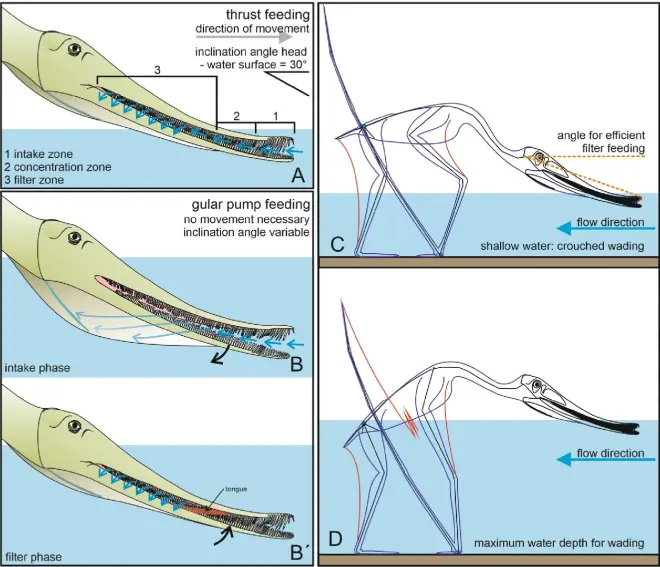
1月21号,期刊《Palaeontologische Zeitschrift》刊登了英国和德国科学家对晚侏罗世德国的滤食性翼龙描述,在研究中,它们命名了出土自托莱特组(Torleite Formation)的毛氏鲸颌翼龙(Balaenognathus maeuseri)。鲸颌翼龙的正模标本NKMB P2011-633是一个几乎完整的骨架,它保留了绝大部分骨骼以及包括翼膜在内的软组织。

鲸颌翼龙属于梳颌翼龙科(Ctenochasmatidae),它的体型较小,翼展只有1.17米,但在梳颌翼龙科里算体型中等。它拥有相当多的独特特征:吻尖形成一个三角形的匙状结构,上面没有牙齿;颌骨牙齿极多,总共至少有480颗;牙齿长而细,齿尖有一个钩状的结构;翼指骨很长,并且肱骨+尺骨与桡骨+第四掌骨的比例也与其它的梳颌翼龙科不同。系统发育分析显示:它与黎明神龙翼龙(Aurorazhdarcho)、剑头翼龙(Gladocephaloideus)、飞龙(Feilongus)、莫干翼龙(Moganopterus)和枪嘴翼龙(Lonchodectes)组成一个单系群,其中鲸颌翼龙处在最基底的位置,但这个支持度并不高,需要进一步研究验证。


鲸颌翼龙独特的形态让科学家相信它很可能是一种滤食性的翼龙,为了佐证这个结论,科学家进行了多方位的分析。首先:鲸颌翼龙的牙齿细长而交错,当它闭嘴时,口腔便会形成一个明显的“筛子”,而齿尖上的小钩可能起到了“卡扣”的作用,辅助上下颌牙齿紧密连接;其次,吻尖的无齿匙状结构扩大了吻前吸水的面积,而鲸颌翼龙上颚的弯曲程度要比下颚更大,这使得它的吻部前端始终是不闭合的,可以在不张口的情况下将水吸入口腔;最后,它拥有发达的下颌关节突以及发达的舌骨器,可以附着更多的肌肉,提高进食能力。
科学家推测:在滤食时,它吻部的前三分之一会完全没入水中,头与水平面呈30°,通过舌骨活动将富含浮游生物和小型动物的水吸进口中,然后再排出,舌骨器和舌头也会控制排水,将这些小型动物留在口中。而有关鲸颌翼龙滤食的方式,科学家提出了几种假设:其一是它在掠过水面或淌水时进行撇滤,但考虑到其头骨结构会在飞行时产生不小的阻力,这种可能性并不大;其二是被动的静止吸食,它可能会选择一些营养丰富的小型水体充当“鳗鱼网”,但考虑到它无法闭合的口腔,这样做的效率似乎并不高;最后一种则是用咽喉充当泵,将营养泵入口中,这点也最被科学家所看好。不管怎么说,鲸颌翼龙展现出了明显且独特的滤食习性,大大地增加了翼龙类的生态多样性。
最古老的引螈型类
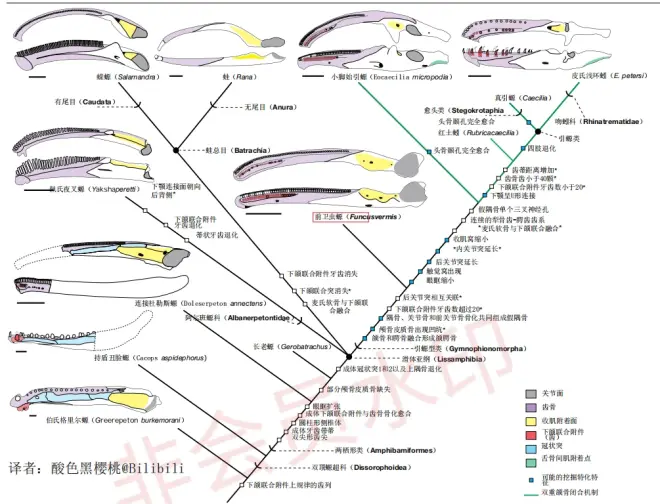
近年来的研究显示:现存的滑体亚纲很可能来自于离片椎类的双顶螈超科(Dissorophoidea),到现在为止,我们只发现了早期的蛙总目(Batrachia)而没有发现原始的引螈类(Gymnophiona)记录。不过在1月25日,期刊《Nature》刊登了目前为止最早的茎群引螈类物种,来自美国的科学家命名了吉氏前卫虫螈(Funcusvermis gilmorei)。前卫虫螈生存于晚三叠世的美国亚利桑那,化石出土于钦迪组(Chinle Formation),年代大致为阿达曼期,正模标本PEFO 43891是部分下颌。

前卫虫螈属于比引螈类更加基干的引螈型类(Gymnophionomorpha),它拥有许多独有的特征:例如下颌联合的小孔以及分隔正中侧和外侧的小凹,颌腭骨与感觉器官无骨学联系、上面的腭齿在内鼻孔的侧缘终止,假隅骨的收肌附着点占据了骨全长的30%、上面有三个三叉神经孔缺乏关节突和内侧突。系统发育分析显示:它是最早且最基干的引螈型类。
作为处于演化关键阶段的物种,前卫虫螈拥有介于双顶螈类和滑体两栖类之间的特征,比如它拥有类似于今天双顶螈类的带蒂牙齿,这一特征在引螈类和蛙总目中依然保留,而在异尾螈目(Allocaudata)的阿尔班螈科(Albanerpetontidae)中消失;还有下颌联合,它的下颌联合并非完全骨化,这与完全骨化的引螈类和蛙总目不同;再比如有关挖掘习性的特化:上颌骨与腭骨融合成颌腭骨、头骨上用于容纳粘液腺的小窝以及用于特化成为管状躯干的背侧扁平的椎弓,这些都是适于挖掘的标志。
前卫虫螈的发现进一步支持了滑体亚纲起源于双顶螈类的假说,并且新的分子钟预测滑体两栖类可能早在中晚密西西比世就已出现,早于以前预测的晚宾夕法尼亚世-早二叠世起源。但由于缺乏这个时期的化石材料,还需要更多的研究来寻找滑体两栖类的起源地的起源时间。
最后,古地理学证据表明:早期的引螈型类可能起源于晚古生代的赤道地区,并在劳亚大陆和冈瓦纳大陆分离前就完成了扩散,但随着板块的漂移,这些适应潮湿热带地区的类群逐渐走向灭绝,剩下的引螈类则继续寻找适宜生存的地区,最终影响了现代无足目的分类情况。

灭绝古猿的牙齿功能学
1月27号,《Journal of Human Evolution》刊登了中国和法国科学家对布氏巨猿(Gigantopithecus blacki)下后犬齿齿冠高和咬合力的估测。布氏巨猿被认为经常取食坚硬食物,因此它可能有着较高的咬合力。为了得出更加全面的结论,科学家选取了16颗布氏巨猿的下颊齿进行分析,并将其与其它的人科物种进行比对。

结果显示:随着牙位的靠后,布氏巨猿的绝对齿冠强度(ACS)和平均牙釉质厚度(AET)都在略微增加,双侧牙釉质底径(BCD)基本保持不变,但众多样本平均下来的平均ACS没有统计学之间的差异,并且都处在一个较高的水平上,这表明布氏巨猿牙齿的抗断裂能力较强,可以抵御很强的外力;而在其它受检测的人科当中,除了鲍氏傍人(Paranthropus boisei),所有物种的磨齿ACS都要低于布氏巨猿,这进一步证明了布氏巨猿要比其它绝大多数人科都适合取食更加坚硬的食物。

对布氏巨猿不同牙齿咬合力的分析显示:它的前磨齿咬合力大于磨齿咬合力;同时,牙尖角(DHA)随着牙位的靠后逐渐变大,牙尖牙釉质厚度(CET)基本无变化。这种情况表明它的前磨齿比磨齿更加尖锐,结合更高的咬合力,布氏巨猿可能主要使用前磨齿进行研磨。在其它受检测的人科物种当中,与它的咬合力最接近的仍然是鲍氏傍人,不过它的咬合力上限却比鲍氏傍人更高。
综合ACS、AET以及咬合力,科学家相信布氏巨猿极有可能确实是一种能够啃食坚硬物体的大型人科物种,它低矮的齿冠、平坦的咬合面以及比腭区更大的前磨齿齿列也提供了形态学上的佐证。但我们目前还不能确定它究竟是以这些坚硬食物为主食还是作为一个机会主义者间接地取食这些食物,参考现生的灵长类,当有充足的营养丰富且易咀嚼的食物存在时,它们并不会大量进食坚硬的食物;同样地,南方古猿和傍人的牙齿微磨损和掉牙也不支持它们将这些坚硬的物体作为常规食物,因此巨猿应该也同样是一个偶尔进食坚硬食物的多面手。后续还需要寻找更多的材料以及对已知的材料进行更加全面的分析来进一步构建这种已灭绝大型人科物种的食谱。


古新世的巨型企鹅
近几年来,新西兰发现了不少尺寸巨大的企鹅目(Sphenisciformes)化石,而在2月8号,期刊《Journal of Paleontology》刊登了美国、新西兰等国科学家对它们的描述,在研究中,它们基于一些材料命名了两个新物种:弗氏巨鸟企鹅(Kumimanu fordycei)以及斯氏矶泳企鹅(Petradyptes stonehousei)。弗氏巨鸟企鹅和斯氏矶泳企鹅都生存于晚古新世的新西兰,化石均出土于摩拉基组(Moeraki Formation),前者的正模标本NMNZ S.47426包含一些颈椎、肩胛乌喙骨、前肢以及后肢,后者的正模标本NMNZ S.47114则是肱骨和部分股骨。

弗氏巨鸟企鹅是巨鸟企鹅属的第二个种,相比于模式种毕氏巨鸟企鹅(Kumimanu biceae),它的体型明显更大,肱骨体更加弯曲、近端扩张程度低。它可能是目前已知最大的企鹅目成员,跟据肱骨长度和肱骨头最大宽度建立的公式可以分别得到最大148kg以及159.7kg的数值,而毕氏巨鸟企鹅的两个值分别为116.1kg和121kg,不难看出尽管存在着一些误差,但弗氏巨鸟企鹅毫无疑问是已知最大企鹅目的有力竞争者。

矶泳企鹅是一个新属,目前仅有斯氏矶泳企鹅这一个种,它与其它古新世企鹅最大的不同就在于它的肱骨体十分扁平,肱骨体中部长轴宽几乎是短轴宽的2倍。相比于巨鸟企鹅属,它肩胛喙骨肌的附着瘢痕几乎与肱骨三叉窝平齐,而后者几乎是向斜后侧延伸。由于化石的不完整性,利用肱骨头最大宽度以及肱骨体中部宽分别计算得到50.3kg和31.2kg的估值,误差较大,但可以看出相比于巨鸟企鹅,矶泳企鹅的体型要明显小一些。

系统发育分析显示:巨鸟企鹅与矶泳企鹅比早古新世新西兰的原始企鹅例如海企鹅(Waimanu)和后海企鹅(Muriwaimanu)更接近冠群,但比潜鸟企鹅(Kupoupou)更加基干。弗氏巨鸟企鹅的演化位置表明了企鹅目在其演化的早期阶段就已经诞生了体型庞大的物种。先前的研究表明,企鹅目很可能起源于古西兰大陆,并在肱骨丛演化完成后扩散到世界各地,而企鹅目早期的体型增大也可能与体温调节有关,并在某种方面促进了它们的扩散。
值得注意的是:巨鸟企鹅和矶泳企鹅的前肢肌肉附着点类似于今天的一些潜水鸟类例如鹈燕和海雀,它们都拥有发达的胸肌和肩胛喙骨肌切迹,可以帮助其在空中上下俯冲以及在水中潜泳,这表明这些早期的茎群企鹅仍然保留了一些祖先的特征,它们不会飞,因此这些曾经对飞行来说非常重要的特征在后期的冠群企鹅中逐步演变为更加适于游泳的形态。也因此,科学家认为这些早期企鹅的游泳能力并不及更加衍化的冠群企鹅,不过这点还需要更多的定量分析。

三叶虫的鳃部清理
从水中高效地提取氧分对早期节肢动物的演化有着至关重要的作用,而如何清理这些频繁使用的呼吸器官也是它们所需要面临的一个难题。2月15号,《Geological Magazine》刊登了中国和美国科学家在寒武纪三叶虫上发现的刚毛化石,在研究中,他们在两个物种上面都发现了刚毛的存在:中寒武世的锯刃拟油栉虫(Olenoides serratus)以及晚奥陶世的伊氏三分节虫(Triarthrus eatoni)。
拟油栉虫的刚毛位于步足,第三到第五肢节的背侧(第二肢节可能也有),刚毛的长度是鳃丝长度的2.5倍左右,这些刚毛通常不位于鳃支的正下方,只在第三肢节有一定的重合,不过当拟油栉虫开始运动,步足进行前后旋运动时,这些刚毛正好可以与鳃支进行接触,完成简单的清扫。不仅如此,科学家测量后发现:刚毛的间距要窄于鳃丝间距,这可能使得每一个鳃丝间隙都可以得到清洗。相比之下,位于肢节腹侧的刺则并不存在清理功能,它们的作用可能还是处理猎物。

不同于拟油栉虫,三分节虫的刚毛位于鳃支轴上,近端轴单体上往往只有一条刚毛,而远端轴单体上会有两条以上刚毛,刚毛的长度达到了鳃丝的四倍。三分节虫这样的刚毛分布让科学家认为它的刚毛是用来梳理相邻鳃支的,并且与拟油栉虫不同,三分节虫的刚毛数量少并且比鳃丝更细,科学家认为:这样的分布模式表明它的一根刚毛需要清理多个鳃丝间隙。但无论是刚毛如何分布,它的长度永远是长于鳃丝的,这非常有利于清理鳃丝间的污染物。
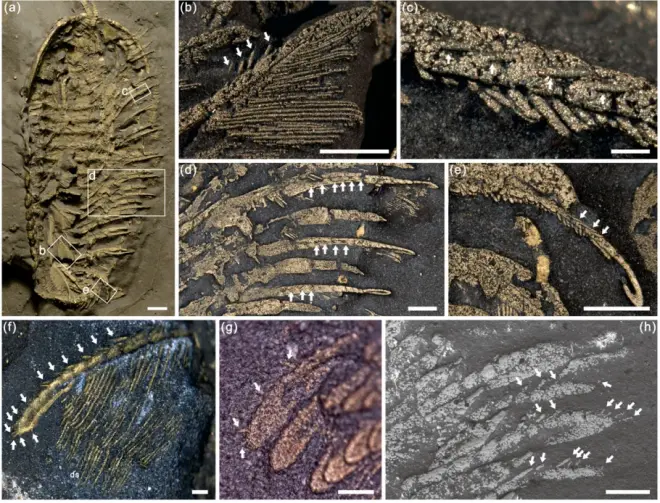
由于三叶虫营底栖生活,并且鳃完全裸露在外,大量的微粒会附着在鳃部或被困在鳃丝之间,因此鳃部的清理对它们来说十分重要。此次发现不仅证明三叶虫会使用刚毛清理鳃支,不同类型的三叶虫之间也拥有不同的清理方式,它们可以通过这样的方式提高呼吸效率,并可能与它们的繁盛存在着些许的联系。最后,科学家还指出:如果三叶虫的蜕皮时间随着个体的发育而逐渐延长,那么鳃部的清理会变得极其重要。

奇特兽脚类手爪的演变
2月16日,期刊《Communications biology》刊登了中国和英国科学家对阿尔瓦雷斯龙超科(Alvarezsauroidea)以及镰刀龙下目(Therizinosauria)前爪的分析研究。在研究中,他们利用系统发育分析以及有限元分析来调查这两个类群前爪的演化,并引入冠龙(Guanlong)和异龙(Allosaurus)作为非手盗龙类的例子进行比对。
研究发现:早期的阿尔瓦雷斯龙类拥有较广的形态空间占据,并且拥有较低的应力水平,而晚期的阿尔瓦雷斯龙类例如临河爪龙(Linhenykus)和单爪龙(Mononykus)则拥有十分短小的爪子,类似于穿山甲。相比之下:除了镰刀龙(Therizinosaurus)本尊以外的镰刀龙类都占据了类似于孤巨懒(Eremotherium)的中低应力位置,镰刀龙本尊则拥有异常高的应力。

进一步的有限元分析显示:所有的镰刀龙类的手爪在刮挖方面表现得都很差,它们更善于穿刺和钩拉,早期的镰刀龙类例如二连龙(Erliansaurus)和铸镰龙(Falcarius)在这两方面表现得比晚期镰刀龙类更好,对它们来说,钩拉树枝和捕捉小猎物或许都是可行的;而晚期的镰刀龙类例如镰刀龙本尊由于手爪高度特化,三种运动方式都会给它的指爪带来损害,同时先前的研究也证实了在镰刀龙类的演化过程中发生的形态变化。据此,科学家认为:镰刀龙类并不会用爪子参与积极的捕猎或进行激烈的搏斗,镰刀龙本尊的爪子甚至可能仅有展示以及装饰的作用,只存在极少量的机械功能。
除了镰刀龙类,阿尔瓦雷斯龙类也得到了十分全面的分析。对单爪龙的有限元分析表明,它的爪子在穿刺、钩拉以及刮挖三个方面都展现出了不错的适应,并且相比于早期的阿尔瓦雷斯龙类例如简手龙(Haplocheirus),它对于挖掘的适应更加明显。因此,晚期的阿尔瓦雷斯龙类确实存在挖掘能力,可以挖掘土堆和数目搜寻昆虫。

经过此次研究,科学家总结并重构了镰刀龙类以及阿尔瓦雷斯龙类内部手爪形态与功能的复杂关系。并且值得注意的是:尽管指爪生前附着着角质爪鞘,但科学家也曾经研究过重建爪鞘后指爪的应力,结果是应力并没有发生明显的改变。因此,在不确定爪鞘的具体形态前,没有明确的证据证明它可以从根本上改变这些奇特手盗龙类恐龙的机械功能。

无角犀亚科的系统发育分析
同样在2月16号,期刊《Frontiers in Ecology and Evolution》刊登了中国和意大利科学家对无角犀亚科(Aceratheriinae)的新系统发育研究,无角犀亚科是一类已经灭亡的曾经广布于欧亚、北美和非洲的犀科(Rhinocerotidae)分支,但由于命名时间相当早、成为过分类垃圾桶加上缺乏足够的形态学和系统发育学研究,它处在一个较尴尬的位置。
新的系统发育分析证明无角犀亚科毫无以为是一个单系群,包含一众基干类群、无角犀族(Aceratheriini)以及远角犀族(Teleoceratini),它被定义为包含巨獠犀(Aprotodon)、远角犀(Teleoceras)以及无角犀(Aceratherium)的最近共同祖先以及所有族裔的分类群。在无角犀亚科中,巨獠犀以及中无角犀(Mesaceratherium)是最基底的成员,二者组成一个支系;曾经被认为属于无角犀族的奇角犀(Alicornops)被认为可能属于远角犀族;不仅如此,系统发育分析还将原无角犀(Protaceratherium)以及图尔卡纳犀(Turkanatherium)排除出无角犀亚科,前者被认为更接近犀亚科(Rhniocerotinae)和板齿犀亚科(Elasmotheriinae),而后者则可能就是一个板齿犀类。

重新建立无角犀亚科的系统发育分析后,科学家也一并提出了这个类群的鉴别特征:体型中等到大型、倾向于演化出较短的头骨、鼻角形态较多样,鼻切迹深、一直延伸到P3-M1处、下方有眶下孔,I1增大或缺失、i2增大,颊齿中等程度特化、唇侧齿带基本退化仅剩部分残留、上前磨齿的舌侧齿带依旧保留,上颊齿的舌侧齿尖收缩或扩张,第五掌骨保留功能、肢骨具备承重能力、远端大而扁。
最后,科学家还在研究的系统发育树中发现了两个尚未被研究过的新支系,第一个就是前面介绍过的“巨獠犀+中无角犀”支系,这一支系生存于晚渐新世和早中新世;另一个支系则是“无鼻角犀(Acerorhinus)+光头犀(Aphelops)+蹄角犀(Hoploaceratherium)”支系,这个类群则生存于中晚中新世。


镰龙类的股骨形态学
2月27日,期刊《The Anatomical Record》刊登了美国科学家对镰龙形态类(Drepanosauromorpha)物种股骨的三维骨学形态分析。由于此前发现的镰龙类股骨多呈现受到挤压的状态,长时间以来对它们的骨学分析一直不完善。为了克服这种问题他们选取了大量来自上三叠统美国钦迪组(Chinle Formation)和多库姆群(Dockum Group)的保存完好的镰龙类股骨进行骨学研究。

研究发现了镰龙类股骨四个与众不同的形态特征:股骨体腹面缺乏内转子,而是存在一个类似于第四转子的结节,这点不同于大多数拥有内转子的早期双孔类,并且第四结节的类似物呈梯形或卵圆形,与主龙形类丘脊形的第四转子有很大的不同;胫骨内侧髁表面平坦,并且内侧面有一个小的亚圆形凹陷以及腓骨髁呈现亚三角形。这样的特征组合在其它的早期双孔类当中是完全没有的。不仅如此,镰龙类的股骨还嵌合了早期双孔类以及主龙形类的特征例如发达的半球形股骨头、股骨头远端结节以及髁间槽等等。

跟据镰龙类股骨的形态,科学家提出了两个假设:其一是镰龙类能够像今天的变色龙一样增大自己大腿的伸展范围,二者的转子承载着类似的功能,但目前阶段的研究还没有找出更多镰龙类与变色龙类功能相似的化石证据;其二是类似于兽孔类和主龙形类的内收型股骨,由于内转子和近腹侧嵴的退化以及第四转子的出现,镰龙类可能做到比变色龙类更加直立的姿势,但由于镰龙类股骨扭曲程度与可以做到直立或半直立的主龙形类有很大的不同,因此镰龙类大概率仍然做不到副矢状面运动。
总体来说,虽然关于镰龙类的研究较为充分,但还需要更多的研究去探索其后肢的姿态,以目前的观点来看,镰龙类的后肢可以做到很多样化的姿势,但整体上的功能可能更类似于今天·的变色龙类。
寒武再无苔藓虫?
不同于在寒武纪爆发式分化的诸多类群,苔藓虫门(Bryozoa)的化石在寒武纪地层中十分罕见,其中一个很重要的原因就是那些疑似属于苔藓虫的化石很难于其它的动物或是藻类进行区分,在这种情况下,被认为属于早期苔藓虫的仅有原蜂巢虫(Protomelission)这一物种。但在3月8号,期刊《Nature》刊登了中国和英国科学家对“原蜂巢虫是寒武纪唯一确定的苔藓虫”这一观点的质疑。
在研究中,他们将发现于下寒武统第三阶小石坝生物群的类似于原蜂巢虫的宏化石并与来自澳洲维拉尔帕的磷矿化的盖氏原蜂巢虫(Protomelission gatehousei)化石进行比对研究。这些来自小石坝的样本呈现出叶形,底部收束,可能用来附着于某些动物的表面;叶状体由巨量的近圆形单体组成,这些单体被薄而坚固的含铁、磷的薄壁包围,并在紧密连接下形成六边形或菱形的结构。将其与维拉尔帕的盖氏原蜂巢虫化石进行比较后,科学家将其归为原蜂巢虫?未定种(Protomelission? sp.)。

科学家将维拉尔帕和小石坝的原蜂巢虫与其它的苔藓虫以及藻类植物进行比较后发现相比于苔藓虫,原蜂巢虫似乎于绒枝藻目(Dasycladales)有着更多的相似之处,例如它与绒枝藻目都有芽状分支、较薄的单体薄壁以及不存在明确且规律的外周小孔(维拉尔帕和小石坝的原蜂巢虫都保留了类似的结构,但这可能与埋藏学有关)。据此,科学家认为原蜂巢虫实际上可能是一类原始的绒枝藻目成员,它的出现时间甚至早于冠群绒枝藻目的分化时间。
原蜂巢虫的存在证明这些底栖的原始藻类在寒武纪的群落发挥着重要的作用,尽管它们在这类生物群当中的分布并不广泛。并且如果原蜂巢虫属于绒枝藻目,那么可以证明早在寒武纪第三期这样的大型藻类已经可以利用外界环境矿化。不过也因此,寒武纪已经不再有确定的苔藓虫类存在,尽管依然存在某些可能被归为苔藓虫的存在,但未来必须需要更多的研究去分析它们的归类。

最早的三叠纪鱼龙?
在以前的认知中,鱼龙形下纲(Ichthyosauromorpha)作为三叠纪蜥形纲的海洋开拓者,在史密斯-斯帕斯灭绝事件以后才下水并很快繁盛起来,支持这一观点的证据是目前发现最早的鱼龙化石都集中在248.8Ma,不过在3月13号,期刊《Current Biology》刊登的通信报告上,来自瑞典和挪威的科学家公布了目前已知最早的鱼龙化石。这个编号为PMO 245.975的样本出土于挪威下三叠统的维京赫格达组(Vikinghøgda Formation),年代大约为250Ma,要早于史密斯-斯帕斯灭绝事件(249.6Ma前后),距离二叠纪末大灭绝(251.9Ma前后)也只有约200万年。

PMO 245.975包含11块椎骨以及一些零碎材料,椎骨呈现双凹形,中央有脊索神经管,关节面背腹拉长,呈六边形;显微CT扫描显示:这些椎骨的椎弓连接面位于腹侧缘并且缺乏连接肋骨的隆起,类似于早期鱼龙超目(Ichthyopterygia)的远端尾椎;骨组织分析显示它内部由松质骨组成,包含一个向四周放射的骨小梁网络,这些是个体成年的特征,并且象征着快速生长、代谢增加的远洋习性。科学家预测,它的体型可以达到3米,是一种适应了远洋的鱼龙超目成员。

新标本的发现证明鱼龙在很早以前就已经游弋在海洋之中,科学家也因此提出了一个极具开创性的观点:鱼龙形下纲早在二叠纪末大灭绝之前就已经下水,而不是在中生代早期作为补充生态空缺的存在出现。以这个推测为大框架,科学家认为鱼龙的早期演化是机会性地进行了生态位的分化,并经历了浅海的两栖动物主导的生态系统以及深水远洋的爬行动物主导的生态系统。不过由于目前缺乏更多的证据,还需要更多的化石材料以及相关研究去验证这个研究的真实性和可靠性。

横贯三洲的迅捷之熊
3月14号,《American Museum Novitates》刊登了中国和美国科学家对一类新种大熊猫亚科(Ailuropodinae)物种的研究描述,在研究了大量来自东亚和北美的化石后,他们命名了一个属于郊熊族(Agriotheriini)的新属:风神熊属(Huracan),这个属下包含许多个种:模式种是师氏风神熊(Huracan schneideri,也就是曾经的师氏郊熊Agriotherium schneideri),来自亨普菲尔期末期(Hh4)并可能延续到早勃朗期的“北美陆地哺乳动物时代”(NALMAs);科氏风神熊(Huracan coffeyi)来自北美Hh3的NALMAs;邱氏风神熊(Huracan qiui)来自中国保德期的“亚洲陆地哺乳动物时代”(ALMA);罗氏风神熊(Huracan roblesi)来自晚中新世-早上新世(MN13)的西班牙。除此之外还有尚待研究的旁遮普“风神熊”?(?“Huracan” punjabiensis)。

风神熊是一种大型的熊类成员,头骨比例大,最大的头骨可以达到515mm,齿列展现出了明确的巨食性,并且作为一个独立属,它拥有许多诊断特征:下颌咬肌前窝存在,I1和I2的后中侧尖发达、后侧尖退化,I3和犬齿发达,以及前磨齿存在、但有时候在除邱氏风神熊以外的几种风神熊中退化等等。而风神熊最明显的就是运动能力的特化,对科氏风神熊的颅后骨骼研究显示:它肱骨上髁不发达、外上髁向近端延伸并与三角肌结节重合,并且肱骨远端的宽度与肱骨长度的比值要比现生的一些熊类都要低、反而更接近一些豹属成员。这些特征表明风神熊的前肢旋转、伸展、屈曲的能力都不强,但相对地,它的行走能力很强,拥有十分不错的行走/奔跑速度,这点也可以被它发达的距骨佐证。


系统发育分析显示:郊熊族目前包含三个属:郊熊属、印度熊属以及风神熊属,它们与今天的大熊猫有许多齿列上的相似之处,但郊熊族展现出了很明显的肉食特化。年代分析显示:邱氏风神熊是最早出现的风神熊属成员,距今大约700万年,并在不久之后演化出了衍化的科氏风神熊和罗氏风神熊,不过欧洲的风神熊并没有延续很久,北美的风神熊则一直延续到了中上新世,而未描述的材料显示亚洲的风神熊一直延续到更新世最早期。
而在郊熊族当中,印度熊是出现时间最早且最基干的类型,但它很可能是一个并系群,因为某些早期的印度熊例如似熊印度熊(Indarctos arctoides)与其它所有的郊熊族成员是姊妹演化支。随后它进入亚洲,并可能在那里演化出了郊熊属和风神熊属,三个属在晚中新世的甘肃和政共存,而在山西保德、云南禄丰和元谋只发现了印度熊。
最后,风神熊和郊熊拥有类似的古生态学,都是善于行走/奔跑的巨食性掠食者,它们作为大熊猫亚科的分支在不同的大洲有不同的分布模式,但它们都是新近纪陆地食肉动物群落的重要组成部分。


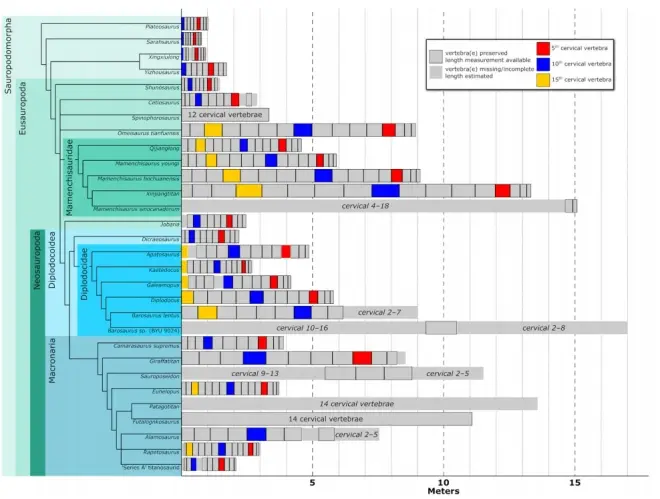
中加马门溪龙的再研究
3月15号,期刊《Journal of Systematic Palaeontology》刊登了中国和英国科学家对著名的中加马门溪龙(Mamenchisaurus sinocanadorum)的再研究。他们重新鉴定了中加马门溪龙的正模标本兼唯一标本IVPP V10603,并重新修订了它的自有衍征:异常延长的外上颌孔,长高比达到了3.7;枢椎腹侧有一深的凹槽、凹槽前方有多个气腔孔以及第三颈椎腹侧靠近正中位置存在发达的、不规则的大量气腔孔。

系统发育分析显示:特征矩阵的选取、不同特征的权重以及不同的演化树建立方式都会很大地影响中加马门溪龙的演化位置,并且马门溪龙属极有可能不是一个单系群,表明对马门溪龙尤其是对马门溪龙属的模式种建设马门溪龙(Mamenchisaurus constructus)进行进一步的分类修订是十分重要的。在某几个演化树中,中加马门溪龙与新疆巨龙(Xinjiangtitan)关系十分亲近,并且二者也都是生存于中晚侏罗世中国西北地区的长颈蜥脚类(新疆巨龙可能比中加马门溪龙更早),不过二者仍然存在许多解剖学的差异,因此并不应当将新疆巨龙草率地归入马门溪龙属。
有趣的是:在研究中,科学家认为巧龙(Bellusaurus)以及大安龙(Daanosaurus)都可能是幼年的马门溪龙类,它们的一些特征代表着它们处在个体发育的早期阶段。

作为一种巨型的马门溪龙类,中加马门溪龙具有相当长的脖子,而在这次的研究中,科学家基于新疆巨龙的枢椎和第三颈椎得出了中加马门溪龙14.4米颈长的估值,要大于基于合川马门溪龙(Mamenchisaurus hochuanensis)的估值(11米),这表明马门溪龙类的颈椎会随着体型正向异速生长;而利用同样拥有较完整颈椎序列的杨氏马门溪龙(Mamenchisaurus youngi)可以粗略估测到15.1米,这使得中加马门溪龙成为了最长脖子的有力竞争者。不仅如此,中加马门溪龙4.2米的颈肋比其它任何一种蜥脚类的颈肋都大。由于其较大的体型和极长的脖子,中加马门溪龙在形态学方面发生了很多变化。其一:它的颈椎骨骼气腔化程度非常高,可以在很大程度上减重;其二:极长的颈肋可以稳定住长脖子的轴向运动,它的颈部运动能力可能因此不及其它的马门溪龙属,但也因此获得了颈部的极端伸长。

食鱼鳄超科演化的新启示?
3月16号,期刊《Journal of Anatomy》刊登了英国科学家对中新世北非的道氏“切喙鳄”(‘Tomistoma’ dowsoni)的神经解剖学研究,并希望借此来进一步探究马来鳄与食鱼鳄之间以及它们早期食鱼鳄类的脑颅神经学演化。他们选取了来自早中新世埃及的标本NHMUK PV R 4769并对其进行脑颅和内耳的重建。

研究发现:道氏“切喙鳄”的脑部结构在许多方面都介于食鱼鳄和马来鳄之间,例如马来鳄的鼻咽管在鼻旁窦后侧分叉,而其它两者分叉的位置基本与鼻旁窦平齐;鼻腺凹集中在吻后端,而其它两者的鼻腺凹分布更靠前;不同位置的大脑扩张程度近似,而其它两者在脑末端扩张最明显。同时,食鱼鳄的嗅区和脑嗅球的连接较平缓,而其它两者嗅区和脑嗅球的连接较锋利。脑容量计算显示:道氏“切喙鳄”的脑容量为23,405立方毫米,介于马来鳄(19,390立方毫米)和食鱼鳄(38,309立方毫米);REQ(爬行动物脑化商)计算得0.86,落在了食鱼鳄的REQ范围内(0.8-2.0)。结合这些特征,我们可以得出道氏“切喙鳄”依然是一类典型的长吻半水生的鳄鱼。


结合此前的系统发育分析以及此次的神经解剖学,道氏“切喙鳄”比起今天的马来鳄更接近食鱼鳄,它可能是一个独立的新物种,目前科学家正在对其分类进行更进一步的分析研究。
历史上切喙鳄类和食鱼鳄类的分类曾存在过很多的争议:分子证据证明二者有着十分亲近的演化关系,但一些形态学研究却将食鱼鳄类归为与切喙鳄类十分遥远的分支。不过随着研究的推进,形态学也发现了足够的证据支持二者的亲缘关系,那些在历史上被归为“切喙鳄”的物种也逐步地得到了清理,而此次对切喙鳄的研究也证明它可能是一个更接近食鱼鳄类的物种。后续的研究将会进一步探讨这些早期食鱼鳄类与现存两个属的分类亲和力以及脑颅神经解剖在食鱼鳄类分类当中的作用。

到此为止,2023第一季度的古生物总结到这里就告一段落了,总体来说,这一季度虽然有着许多可圈可点的新研究,不过对于部分类群来说有关的研究却并不是那么的惊艳。而我所能做的便是找寻众多研究中的亮点,并呈现给大家,希望大家能够喜欢。
