
从头开始的蛋白质逻辑门设计
引言
蛋白质与蛋白质之间的相互作用在细胞决策中无处不在,控制它们在合成生物学中将变得越来越重要。虽然蛋白质相互作用是天然生物电路的核心,但创建新逻辑电路的工作主要集中在 DNA、转录或 RNA层面的控制。最近,通过重新连接原生信号通路、用盘绕线圈将蛋白质连接在一起或创建蛋白酶级联,产生了基于蛋白质的电路;然而,这些电路都是由有限的构件库构建的,这阻碍了它们的可扩展性。从头开始设计基于蛋白质的逻辑门,调节任意的蛋白质-蛋白质相互作用,可以为细胞内外新的基于蛋白质的控制系统打开大门。
逻辑门设计原则
原则上说,利用一组异源二聚体分子从头设计各种逻辑门是可能的。例如,给定假想的异源二聚体对 A:A′、B:B′ 和 C:C′,通过基因融合 A′和 B,以及 B′和 C,就能构建出调节 A 与 C′关联的 AND 门:只有在 A′-B和B′-C同时存在的情况下才会发生关联(此处及下文中的":"表示非共价相互作用,"-"表示通过柔性连接体进行的基因融合)。在构建这种关联逻辑门时,需要具备几个构件特性。
首先,应该有许多相互正交的异二聚体对,这样逻辑门的复杂性就不会受到单个元素数量的限制。
其次,构件应具有模块化和相似的结构,这样在构建逻辑门时就不必考虑构件形状和其他特性的差异。
第三,单个构件应能以不同的可调亲和力与多个伙伴结合,从而通过破坏已有的低亲和力相互作用来执行否定操作。
第四,相互作用应该是合作性的,从而使门的激活对输入中的化学计量失衡不敏感。例如,在上述 AND 门中,如果相互作用不合作,那么大量过剩的 A′-B 将使平衡趋向于部分组装的复合物(A′-B 与 A 或 B′-C,但不是两者),这将限制门的激活。

图表说明概述了设计基于蛋白质的协同逻辑门研究的主要方面:(A) A:A'异源二聚体结构单元的骨架结构及其氢键网络示意图。(B) 介绍描述诱导二聚系统的热力学循环。(C) 模拟展示了诱导二聚系统在热力学平衡下的行为。在最终形成的三聚复合物中,合作性的影响非常明显,当二聚体蛋白过量时,三聚复合物的形成就会减少。(D) 通过圆二色性(CD)监测 6 氨基酸和 12 氨基酸连接体设计的平衡变性实验。(E)带有 6 个氨基酸连接体的 1'-2' 的小角 X 射线散射(SAXS)实验曲线,与 1:1' 异源二聚体的计算曲线相匹配。(F)带有六位残基连接体的诱导二聚系统示意图,以及随后面板中的实验结果。(G)在 1 存在和不存在的情况下,2 对 1'-2' 的本征质谱滴定。(H) 1'-2' 与 1 和 2 的 nMS 滴定。(I) 在酵母中测试诱导二聚化系统的示意图及体内结果。(K)两输入 AND 门示意图及 nMS 滴定结果。(M)三输入 AND 门示意图,以及 nMS 滴定结果。
作者探索了利用全新设计的具有氢键网络介导的特异性的异二聚体来设计满足上述所有四项标准的逻辑门的可能性。具有氢键网络介导特异性的相互正交设计异源二聚体(DHD,以下用数字表示,如 1 和 1′ 构成一对同源物;表 S1)(如图 1A)可用于构建逻辑门,满足条件 1(正交性)。异源二聚体界面都具有相同的四螺旋束拓扑结构(图 1A),满足条件 2(模块化)。共享的相互作用界面允许配对之间进行有限的交叉对话,从而形成了结合亲和力的层次结构,满足了条件 3(多种结合特异性)。受自然界合作系统的启发,作者试图通过构建单体融合体(上例中的 A′-B 和 B′-C)来实现条件 4(合作性),即在融合体内埋藏(与 A 和 C′的)相互作用界面。暴露这些埋藏界面所需的自由能将阻碍栅极活化,因此作者推断,可以对系统进行调整,使两个伙伴的结合能之和足以克服这一障碍,而不是任何一个伙伴单独的结合能之和,从而确保合作栅极活化。如果条件 2(模块化)成立,那么确保合作性的单一方案原则上可以适用于多种栅极配置。
如果结合是非合作的,当 A′-B过量时,三聚体复合物的形成就会减少。相反,如果是合作结合,则三聚体复合物的形成对二聚体过量不那么敏感。假设二聚体(A′-B)的折叠四螺旋束状状态由于埋藏的疏水相互作用表面而反对与 A 或 B′结合。随后,作者测试了连接 A′和 B 的不同长度的柔性连接体。这些构建体保持稳定。原位质谱法(nMS)用于研究体外的合作性,结果表明,加入 1 后,2 与 1′-2′ 之间的结合显著增加。此外,即使在 1′-2′ 过量 6 倍的情况下,结合仍然很牢固。利用基于酵母的检测方法,活细胞中的合作性也得到了证实。
更复杂的逻辑门设计
进一步的实验探索了更复杂的逻辑门,如双输入 AND 门和三输入 AND 门,结果表明即使在这些更复杂的配置中,合作结合仍然有效。
研究随后扩展到设计一系列双输入合作诱导蛋白异二聚体(CIPHR)逻辑门。这些逻辑门涉及设计的异源二聚体蛋白(DHD)的模块化组合及其与效应蛋白的融合,以根据输入控制共聚或解离。这项研究证明了在体外和活细胞中利用全新设计的蛋白质相互作用设计和实施基于合作蛋白质的逻辑门的可行性,并探索了基于这些原理构建更复杂逻辑门的方法。

图中展示了基于合作蛋白的双输入逻辑门(CIPHR 门)的构造和功能:(A) CIPHR 逻辑门由设计的异源二聚体蛋白质(DHD)构建而成,输入为单体或共价连接的单体。门利用设计的同源相互作用或观察到的结合亲和力层次。(B)利用正交 DHD 相互作用构建的双输入 AND 门。(C) 基于正交 DHD 相互作用的双输入 OR 门。(D) 利用多特异性和竞争性蛋白质结合构建的 NOT 门。(E) 基于多特异性和竞争性蛋白质结合的 NOR 门。(F) 利用多特异性和竞争性蛋白质结合构建的 XNOR 门。(G) 基于多特异性和竞争性蛋白质结合的 NAND 逻辑门。
对于每个逻辑门,黑点代表经背景生长校正的单个酵母双杂交(Y2H)生长测量值,其平均值用绿条表示。灰色方框表示使用的 DHD 对,蓝色方框表示亲和力梯度。输入状态和预期输出分别用中间和右边图块中的 "0 "和 "1 "表示。图中展示了这些基于合作蛋白的逻辑门在各种逻辑运算中的通用性和功能性。
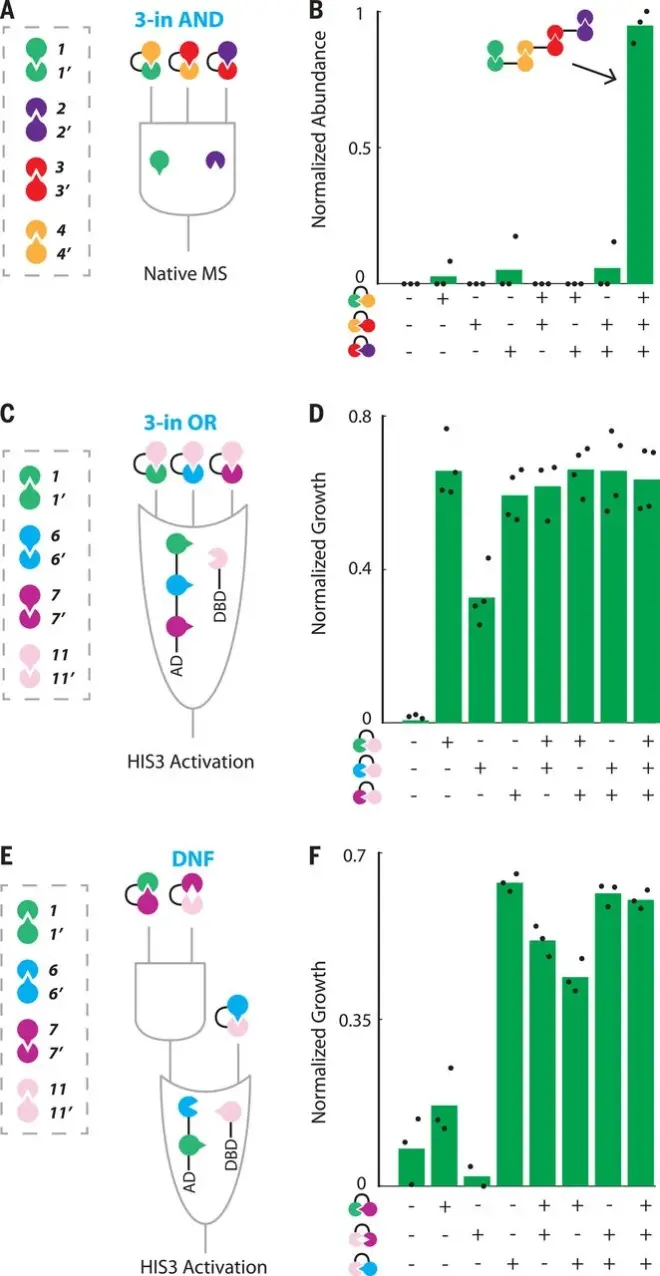
本图展示了基于蛋白质的三输入合作逻辑门(CIPHR 门):(A) 一个三输入 AND 逻辑门的示意图,其中单体 1 和 2 通过输入 1′-4′、4-3′ 和 3-2′ 结合在一起。(B) nMS 结果证实,只有在所有三个输入都存在的情况下,三输入 AND 门才能被正确激活。(C) 三输入 OR 门的示意图,其中任何一个输入(11-1、11-6′或 11-7′)都通过 1′、6 或 7 将 AD 连接到 DBD。(D)酵母-两系杂交(Y2H)结果表明,任何一个输入端都能激活三输入 OR 门。(E) 分立正则表达式(DNF)门的示意图。(F) 证明 DNF 逻辑门正确激活的 Y2H 结果。每个门的单个测量值以黑点表示,平均值以绿条表示。灰色方框表示门中使用的 DHD 对。
图中展示了基于设计的蛋白质相互作用成功构建和运行的三输入 CIPHR 逻辑门。研究利用设计的异源二聚体蛋白(DHD)构建了各种基于合作蛋白的两输入和三输入逻辑门(CIPHR 门),并评估了它们的功能:
通过将特定输入分别与 Gal4 激活结构域(AD)和 DNA 结合结构域(DBD)融合,创建了 AND 和 OR 逻辑门,只有当两个输入同时存在时,逻辑门才会生长。
其他逻辑门(NOT、NOR、XNOR 和 NAND)是根据观察到的 DHDs 结合亲和力层次,利用多特异性和竞争性蛋白质结合原理构建的。
利用原生质谱(nMS)和酵母-双杂交(Y2H)试验对三输入门(AND、OR 和 CIPHR-双结正则表达式)进行了表征,证实了它们的设计行为。
利用 NanoBiT 分裂荧光素酶系统根据输入状态控制荧光素酶的活性,证明了 CIPHR 逻辑门可用于重组分裂酶的活性。
研究人员探索了 CIPHR 与 TALE-KRAB 系统的结合,以实现可编程治疗应用,从而控制 T 细胞中免疫检查点基因的表达。
CIPHR 门的系统化设计利用了从头开始的蛋白质设计,创造出了多功能的功能逻辑电路,可以与各种蛋白质致动域连接,在不同的环境中提供了潜在的应用。这项研究强调了从头蛋白质设计与 nMS 等实验技术之间的协同作用,并展示了从头蛋白质设计接近天然蛋白质组装复杂性的能力。CIPHR 门为多功能信号处理和蛋白质-蛋白质相互作用控制提供了一个平台,对包括合成生物学和治疗应用在内的各个领域都有潜在影响。参考资料:
Chen Z, Kibler R D, Hunt A, et al. De novo design of protein logic gates[J]. Science, 2020, 368(6486): 78-84.
版权信息
本文系AIDD Pro接受的外部投稿,文中所述观点仅代表作者本人观点,不代表AIDD Pro平台,如您发现发布内容有任何版权侵扰或者其他信息错误解读,请及时联系AIDD Pro (请添加微信号sixiali_fox59)进行删改处理。
原创内容未经授权,禁止转载至其他平台。有问题可发邮件至sixiali@stonewise.cn
