
细胞生物学复习
第一章
细胞学说建立者:施莱登、施旺
细胞类型:真核细胞、原核细胞、古核细胞
原核细胞与真核细胞基本特征的比较:



原代细胞:机体取出后立即培养的细胞。
传代细胞:适应在体外培养条件下持续传代培养的细胞称为传代细胞。
细胞系:细胞系(cell line)指原代细胞培养物经首次传代成功后所繁殖的细胞群体。 也
指可长期连续传代的培养细胞。
细胞株:通过选择法或克隆形成法从原代培养物或细胞系中获得具有特殊性质或标志物
的培养物称为细胞株(Cell Strain)。
第二章
光学显微镜原理:光学显微镜主要由目镜、物镜、载物台和反光镜组成。目镜和物镜都
是凸透镜,焦距不同。物镜的凸透镜焦距小于目镜的凸透镜的焦距。物镜相当于投影仪
的镜头,物体通过物镜成倒立、放大的实像。目镜相当于普通的放大镜,该实像又通过
目镜成正立、放大的虚像。
电子显微镜原理:在真空条件下,电子束经高压加速后,穿透样品时形成散射电子和透
射电子,它们在电磁透镜的作用下在荧光屏上成像。电子束投射到样品时,可随组织构
成成分的密度不同而发生相应的电子发射,如电子束投射到质量大的结构时,电子被散
射的多,因此投射到荧光屏上的电子少而呈暗像,电子照片上则呈黑色。主要优点:分
辨率高,可用来观察组织和细胞内部的超微结构以及微生物和生物大分子的全貌。
荧光染料:

第三章 细胞质膜
细胞膜定义:细胞膜(cell membrane)又称质膜(plasma membrane)是指围绕在细胞
最外层,由脂质和蛋白质组成的生物膜。
细胞膜基本功能:是细胞结构上的边界;维持细胞相对稳定的内环境;细胞与环境之间、
细胞与细胞间进行物质、能量交换场所;信息传递等。
流动镶嵌模型结构要点:(1)生物膜由磷脂双分子层和蛋白质构成,磷脂极性头部朝向
水相;非极性尾部聚集在一起,以避开水;(2)蛋白质以不同方式镶嵌于膜质中,或通
过氢键、离子键等方式结合于表面;(3)具有流动性,并在生命活动中处于动态变化
中;(4)结构具有不对称性
脂筏模型:脂筏是以甘油磷脂的生物膜上,胆固醇和鞘脂形成相对有序的脂相,如同漂
浮在脂双层上的“筏”一样,载着具有生物功能的膜蛋白。
流动镶嵌模型结构要点:
膜脂的运动方式:(1)沿膜平面的侧向运动(基本运动方式),其扩散系数为 10
-8cm2/s;
(2)脂分子围绕轴心的自旋运动;(3)脂分子尾部的摆动;(4)双层脂分子之间的
翻转运动(flip-flop)
脂质体及其意义:
脂质体:脂质体是根据磷脂分子可在水相中形成稳定的脂双层膜的现象而制备的人工膜。
意义:研究膜脂与膜蛋白及其生物学性质; 脂质体中裹入 DNA 可用于基因转移,在
临床治疗中,脂质体作为药物或酶等载体。
外在蛋白:非共价键与膜表面结合,与磷脂头部发生作用。
内在蛋白(整合膜蛋白):蛋白质显极性,膜内蛋白质的极性区突向膜表面,非极性部
分则埋在双层内部。
脂质锚定蛋白:(1)与脂肪酸结合;(2)通过异戊二烯单位结合到 Cys 残基上;(3)
通过糖脂锚定
细胞质膜的不对称性:是指膜蛋白和膜脂等在生物膜上呈现不对称分布,糖脂、糖蛋白
的糖链均位于质膜的外侧。其次是膜蛋白分布的不对称性,有的镶在膜的表面,有的嵌
入或横跨脂双分子层。
膜脂的流动性及其证明方式:膜脂的流动性:是脂分子的侧向运动。证明方式:荧光漂
白恢复。
膜骨架:指细胞质膜下与膜蛋白相连的由纤维蛋白组成的网架结构,它参与维持细胞质
膜的形状并协助质膜完成多种生理功能。
红细胞血影(Blood ghost)RBC:经低渗处理,质膜破裂,释放出 Hb 和其它可溶性蛋白,
RBC 仍保持原来基本形状和大小。
第四章 物质的跨膜运输
物质通过细胞膜的转运主要有三种途径及其特征:
物质通过细胞膜的转运主要有三种途径:①被动运输 ②主动运输 ③胞吞与胞吐作用。
被动运输:
1. 简单扩散(simple diffusion):小分子物质以热自由运动的方式顺着电化学梯度或浓
度梯度直接通过脂双层进出细胞,既不需要细胞供能,也不需要膜转运蛋白的协助。
特点:顺浓度或电化学梯度;不耗能
2. 协助扩散:在膜转运蛋白的协助下,物质从高电化学势或高浓度一侧向低电化学势
或低浓度一侧的跨膜运输形式,又称协助扩散 (facilitated diffusion )。
特点:顺浓度或电化学梯度;需要载体;不耗能
主动运输:
由载体蛋白所介导的物质逆着电化学梯度或浓度梯度进行跨膜运输的方式。它是一种需
要消耗能量的物质跨膜运输过程。
特点:逆浓度或电化学梯度;耗能
类型:根据所需能量来源不同,分为三种方式:①ATP-driven;②Couple transporter;③
Light-Driven
胞吞与胞吐作用:
胞吞作用(endocytosis ):通过质膜内陷形成膜泡,将细胞外或细胞质膜表面的物质
包裹到膜泡内并转运到细胞内,包括吞噬作用和胞饮作用。
胞吐作用(Exocytosis):与细胞的胞吞作用相反,胞吐作用是将细胞内的分泌泡或其他
某些膜泡中的物质通过细胞质膜运出细胞的过程。
膜转运蛋白、载体蛋白和通道蛋白,类型及特点:
膜转运蛋白:也叫膜运输蛋白。能选择性地使非自由扩散的小分子物质透过质膜。
载体蛋白:通过蛋白质构象变化使得溶质分子穿过细胞质膜,介导主动或被动运输。
功能:普遍存在、多次胯膜,只与特定溶质结合,具有选择性、类似酶的动力学特征,
但不修饰转运的物质. 通道蛋白:形成亲水性通道贯穿质膜,介导顺电化学梯度的被动运输。通道蛋白所介导
的被动运输不需要与溶质分子结合,横跨膜形成亲水通道,允许适宜大小的分子与带电
荷的离子通过。三种类型:离子通道、孔蛋白、水孔蛋白。
葡萄糖的转运方式:葡萄糖在体内的转运以易化扩散的方式进行,顺着浓度梯度进行,
不耗能,但需要特殊的载体(葡萄糖转运蛋白);但是在小肠上皮细胞以主动转运进行
吸收。
葡萄糖转运蛋白:糖转运蛋白家族成员之一,GLUT1 几乎存在于人体每一个细胞中,是
红细胞和血脑屏障等上皮细胞的主要葡萄糖转运蛋白,对于维持血糖浓度的稳定和大脑
供能起关键作用。

离子通道的类型及特征:
离子通道类型为三种类型的门控作用:电压门控、配体门控、应力激活。
离子通道三个特点:(1)转运速率高 107-108/s per channel,动力为跨膜电化学梯度;
(2)没有饱和值;(3)非连续开放,开和关两种构象,而是门控的,类似 All or No。
Na-K 泵:
特点:钠钾泵-具有 ATPase(也称为 F0F1-ATP 合酶)活性,故 Na+-K+-ATPase,只位于动
物细胞质膜上。
结构:四聚体
机制:Na+依赖的磷酸化,K+依赖的去磷酸化,导致构象变化,每消耗 1 个 ATP,泵出 3
个 Na+泵进 2 个 K+,受 Ouabain 抑制。
ATP 驱动泵的主要类型:

P 型离子泵:2 个独立的 a-催化亚基,ATP 结合位点,2 个,起调节作用 b 亚基组成,依
赖 a 亚基磷酸化与去磷酸化过程,导致蛋白质构象改变。磷酸化中间体。
V 型质子泵:通过细胞器和液泡的膜主动运输 H+。它们存在于溶酶体、分泌颗粒和植物
细胞液泡中,也在多种细胞(肾小管)的质膜中被发现。保持细胞质 pH 恒定。
F 型质子泵:位于线粒体内膜,叶绿体类囊体膜、原核生物质膜上,实际上是 H+-ATP
合成酶(F1F0-ATPase)
ABC 超家族:ABC 超家族( ATP binding cassette super-family)又叫 ABC 转运蛋白,是一
类利用 ATP 水解释放的能量将糖、氨基酸、磷脂、胆固醇、肽和其它多种小分子物质进
行跨膜转运的 ATP 驱动泵。
协同转运及其类型:
协同转运:两种溶质协同跨膜运输的过程,是主动运输的一种,间接消耗 ATP。
类型:同向转运;反向转运
同向转运:是物质运输方向与离子转移方向相同,如小肠上皮细胞和肾小管上皮细胞吸
收葡萄糖或氨基酸等有机物,就是伴随 Na+从细胞外流入细胞内完成的。
反向转运:是指物质跨膜转运方向与离子转移方向相反,如动物细胞通过 Na+驱动的
Na+-H+对向运输的方式来转运以调节细胞内的 pH。
第五章 细胞质基质与细胞内膜系统
细胞质基质:在真核细胞的细胞质中,除去可分辨的细胞器以外的胶状物质,占据着细
胞质膜内和细胞核膜外的细胞内空间,也叫胞质溶胶 cytosol。
细胞质基质功能:
(1)完成各种中间代谢过程,如糖酵解过程、磷酸戊糖途径、糖醛酸途径等。
(2)蛋白质、脂肪酸合成场所。
(3)与细胞质骨架相关的功能,维持细胞形态、细胞运动、胞内物质运输及能量传递,
为细胞基质其它组分及细胞器提供锚定位点,使复杂代谢途径有序进行。
(4)细胞内的膜相把细胞质基质区室化,并通过质膜或细胞器膜上的膜转运蛋白维持
细胞内外的跨膜电化学梯度,维持细胞稳态。
(5)蛋白质的修饰、控制蛋白质寿命、蛋白质选择性的降解和错误蛋白质重新折叠等。
细胞质基质与蛋白质修饰和降解的关系:
在细胞质基质中,存在泛素-蛋白酶体所介导的蛋白质降解途径,其生物学功能包括—
蛋白质质量监控、影响细胞代谢、信号转导和受体调整、免疫反应、细胞周期、转录调
节和 DNA 损伤修复等。
蛋白酶体:真核细胞质基质内降解蛋白质的大分子复合体,约 50 种蛋白亚基构成,包
含一个 20S 的催化亚基和 19S 的调节亚基,富含 ATP 依赖的蛋白酶活性,可通过泛素化
途径特异性的降解细胞内蛋白质。
泛素:真核细胞中普遍存在的具有热稳定性的高度保守的小分子蛋白(76 个氨基酸残
基),通过其 C 端的 Gly 残基与要被降解的不稳定的或错误折叠的靶蛋白质的 Lys 残基
的ε-氨基形成异肽键(isopeptide bond)而共价连接,并指引该靶蛋白质运到蛋白酶体
中进行降解。
分子伴侣:是一种与其他多肽或蛋白质结合的蛋白质,协助细胞内蛋白质合成、折叠、
装配与分选。
细胞内膜系统:是指细胞内在结构、功能及发生上相关的由膜包绕形成的细胞器或细胞
结构,包括内质网、高尔基体、溶酶体、胞内体、分泌泡等。
内质网结构与功能:
内质网 Endoplasmic (ER):真核细胞内由封闭的管状或扁平囊状膜系统及其包被的腔形成
互相沟通的三维网络结构。占膜系统一半左右,占体积 10%。形态、结构、功能差异—
—发育、细胞周期、生理状况等而异。

内质网应激反应定义及类型:
内质网应激反应(ERS):当某些因素致使内质网生理功能紊乱、 Ca 2+稳态失衡、未
折叠或错误折叠蛋白质在 ER 腔内超量积累,便会激活一些相关的信号通路,引发内质
网应激反应,从而应对环境变化,并恢复 ER 正常状态。
ERS 反应类型:
(1)UPR 未折叠蛋白质应答反应(UPRER):错误折叠或未折叠蛋白过多积累引起,分子
伴侣蛋白表达上调;
(2)EOR (内质网超负荷应答反应):正确折叠蛋白和膜蛋白异常堆积引起;
(3)固醇调节级联反应(sterol regulatory element reaction):胆固醇损耗所致,SREBP
(固醇调控元件结合蛋白)介导;
(4)启动凋亡程序:内质网功能持续紊乱所致。
高尔基体组成、结构和功能:
高尔基体:由大小不一、形态多变的囊泡组成,1898 年由意大利医生 C. Golgi 利用镀银
染色发现,但确证其存在用去近 50 年,电子显微镜发明以后。
结构:电镜下高尔基体结构是由扁平膜囊和大小不等的囊泡构成。
主要功能: ER 合成的蛋白加工、分类、包装和运输到细胞特定部位或分泌到胞外;运
输一部分 ER 合成脂类物质;为细胞内糖类合成工厂。
高尔基体蛋白质分选的三条途径:
(1)溶酶体酶的包装与分选途径(mannose-6-P 标记在 Gi 完成)
(2)可调节型分泌途径;
(3)组成型分泌途径。
细胞内膜系统的标志性酶:
(1)葡萄糖-6-磷酸酶被视为内质网膜的标志酶。
(2)糖基转移酶是高尔基复合体的标志酶。
(3)酸性磷酸酶是溶酶体的标志酶。
(4)过氧化氢酶可视为过氧化物酶体的标志酶。
溶酶体类型、标志酶:
溶酶体是一种异质性的细胞器,指不同的溶酶体的形态大小,乃至其中所含水解酶的种
类都可能有很大的不同。
类型:按生理功能不同阶段划分不同阶段
(1)初级溶酶体(primary lysosome):球形,d=0.2-0.5 μm,内容物均一,不含明显颗
粒物质,外面一层脂蛋白膜围绕,厚 7.5nm,含酸性水解酶如蛋白酶等。
(2)次级溶酶体(secondary lysosome):初级溶酶体与细胞内自噬泡、异噬泡、胞饮
泡或吞噬泡融合形成的复合体,前者谓之自噬溶酶(autophagolysosome),后者谓之异
噬溶酶体(phagolysosome)。
(3)而包含未被消化的物质残存在溶酶体中形成的残余谓之残余体(residual body)或
后溶酶体。

过氧化物酶体及其功能:又称微体,单层膜围绕的含有一种或几种氧化酶类的细胞器,
能够利用分子氧氧化有机物。1954 年由 Rhodin 发现。但到 1970 年代才被看做是与溶酶
体不同的细胞器。
过氧化物酶体的功能:
1、动物细胞(肝细胞或肾细胞)中的功能:
过氧化物酶体可氧化分解血液中的有毒成分,起到解毒作用。乙醇氧化为乙醛
过氧化物酶体中常含有两种酶:依赖于黄素(FAD)的氧化酶,其作用是将底物氧化形
成 H2O2;过氧化氢酶,作用是将 H2O2 分解,形成水和氧气。
2、过氧化物酶体分解脂肪酸等高能分子向细胞直接提供热能。注意:热能与 ATP 能量。
3、在植物细胞中的功能:
(1)在绿色植物叶肉细胞中,它催化 CO2 固定反应副产物的氧化,即所谓光呼吸反应,
即 Calvin cycle 的副产物乙醇酸盐转化为甘油酸。
(2)乙醛酸循环的反应,在种子萌发过程中,过氧化物酶体降解储存的脂肪酸→乙酰
辅酶 A→琥珀酸→葡萄糖。
第六章 蛋白质分选及其功能
信号假说:该假说认为蛋白质首先在细胞质基质游离核糖体上起始合成,当多肽链延伸
至 80 个氨基酸左右后,N 端的信号序列与信号识别颗粒结合,使肽链延伸暂停,防止
新生肽 N 端损伤和成熟前折叠;直至信号识别颗粒与内质网膜上的停泊蛋白(SRP 受体)
结合,返回重复使用,肽链又开始延伸;信号肽将肽链引入内质网膜,内质网腔面的信
号肽酶切除信号肽,肽链继续延伸至完全合成。
信号肽:位于蛋白质 N 端一段由 16-26 个氨基酸残基组成的序列,包含疏水核心区、信
号肽的 C 端和 N 端三部分,是指导分泌性蛋白在 ER 上合成的关键因子。
信号识别颗粒:是存在于细胞质基质中的核糖核蛋白复合体,由 6 种不同蛋白质和一个
7S 小 RNA 分子构成,含有信号肽识别结构域和核糖体结合结构域。
导肽:线粒体、叶绿体和过氧化物酶体的蛋白质的信号序列特称为导肽(leader peptide)
蛋白质分选:在细胞质基质或 RER 合成的蛋白质,依赖蛋白质分子中的分选信号,从而
到达靶细胞器或细胞表面,以发挥其生理功能,这一过程称蛋白质分选(protein sorting)
或者叫靶向转运。
SRP 活性位点:三个活性位点,即(1)信号肽结合位点(2)核糖体识别结合位点,可
阻止翻译(3)ER 膜结合位点。
信号识别颗粒的受体(又称停泊蛋白,docking protein,DP):DP 是内质网膜的整合蛋
白,由α和β亚基组成,可特异地与 SRP 结合。α亚基可结合 GTP,故为 GTP 结合蛋白。
真核细胞蛋白质分选的主要途径与类型:
总体上有两条途径:翻译后途径、共翻译途径
(1)翻译后途径:既可以是 mt, ct, 过氧化物酶体,细胞核蛋白,也可是细胞质基质驻
留蛋白和支架蛋白等,先在细胞质基质游离核糖体上完成多肽链合成,再转运到各细胞
器;
(2)共翻译转运途径:ER,Golgi,溶酶体,质膜,分泌蛋白等,先在游离核糖体上起
始合成后,由信号肽导引转移到 RER,经转运膜泡运至 Golgi 进行加工包装,然后再转
运至溶酶体、质膜或分泌至胞外。
按照分选转运机制,蛋白质转运四种类型:
(1)蛋白质的跨膜转运(transmembrane transport):指共翻译转运途径中蛋白质边合
成边转运进入内质网腔或插入内质网膜(驻留);另指翻译后转运途径中蛋白质在合成
后依不同机制转运到线粒体、叶绿体和过氧化物酶体等细胞器。
(2)膜泡运输(vesicular transport):蛋白质被不同类型的转运膜泡从糙面内质网合成
部位转运至高尔基体进而再分选转移至细胞的不同部位,其中涉及供体膜出芽形成不同
的转运膜泡、膜泡运输和转运膜泡与靶膜的融合等过程。
(3)选择性门控转运(gated transport):在游离核糖体上合成的蛋白质通过核孔复合
体在核-质间双向选择性地完成核输入或核输出。
(4)细胞质基质中蛋白质的转运:蛋白质在细胞质基质中的转运显然与细胞骨架系统
密切相关。
膜泡运输及其特点:
膜泡运输:是蛋白分选运输的特有方式,指蛋白质通过不同类型的转运小泡从 RER 合成
部位转运至 Golgi,进而分选转运至细胞的不同部位。
特点:① 转运过程涉及蛋白质修饰加工和组装;②涉及不同膜泡靶向运输及其调控,如
不同的运输小泡的定向转运、膜泡出芽以及融合的过程。在 TGN 形成不同的转运泡及分
选运输工作。③胞吞作用同样涉及复杂的膜泡运输过程
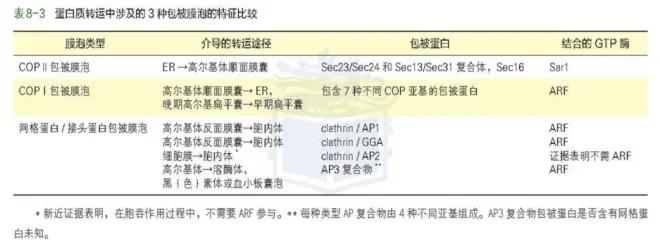
转运膜泡类型:
根据转运膜泡表面包被蛋白的不同,主要分 3 种不同类型:
(1)COPⅡ包被膜泡,介导从内质网向高尔基体的运输,顺向转运。
(2)COPⅠ包被膜泡,负责将蛋白从高尔基体返回内质网,即逆向转运。
(3)网格蛋白/接头蛋白包被膜泡,介导几种蛋白质分选途径,如从高尔基体 TGN 向内
体、溶酶体的分选和运输,还负责胞吞途径从细胞表面至内体的运输。
COPⅡ与 COPⅠ转运小泡运输的类型,衔接蛋白。
COPⅡ运输类型:膜结合蛋白、可溶性蛋白(需要膜受体识别),从内质网至高尔基体
顺面膜囊。
COPⅠ运输类型:(1)从高尔基体反面膜囊到顺面膜囊;(2)从高尔基体顺面管网状
区到内质网的膜泡转运。
网格蛋白/接头蛋白运输类型:(1)网格蛋白/接头蛋白包被膜泡介导蛋白质从高尔基
体 TGN 向胞内体、溶酶体、黑素体、分泌泡和植物细胞液泡的运输;
(2)质膜受体介导的胞吞作用,从细胞表面运往胞内体转而到溶酶体的运输。
第七章 线粒体和叶绿体
线粒体、叶绿体的超微结构:
线粒体的超微结构:
线粒体是由外膜、内膜、膜间隙和基质四部分构成
◆外膜(outer membrane):含孔蛋白(porin),通透性较高。
标志酶:单胺氧化酶
功能:肾上腺素氧化;膜磷脂的合成; 脂肪酸链延伸;色氨酸降解等。
◆内膜(inner membrane):高度不通透性,向内折叠形成嵴(cristae)。含有与能量
转换相关的蛋白。
标志酶:细胞色素氧化酶
功能:电子传递,氧化磷酸化,代谢物质运输 。
◆膜间隙(intermembrane space):含许多可溶性酶、底物及辅助因子。
标志酶:腺苷酸激酶
功能:AMP 的磷酸化,产生 ADP。
◆基质(matrix):含三羧酸循环酶系、线粒体基因表达酶系等以及线粒体 DNA, RNA,
核糖体。
标志酶:苹果酸脱氢酶
功能:丙酮酸氧化,TCA 循环,脂肪的β氧化,DNA 复制,RNA 合成,蛋白质合成

叶绿体的超微结构:
叶绿体是由叶绿体膜、类囊体和基质三部分构成。
◆叶绿体膜:三个层次:外膜、内膜。
外膜与内膜之间是膜间隙。
外膜通透性大,含有孔蛋白
内膜通透性低,含有很多转运蛋白,如磷酸交换载体、二羧酸交换载体
◆类囊体:叶绿体内部由内膜发展而来的封闭扁平膜囊,其内空腔谓之类囊体腔。
类囊体有序叠置形成,种类有:基粒类囊体、基质类囊体
基粒类囊体:在某些部位,许多圆饼状的类囊体叠成垛,称为基粒。组成基粒的类囊体
称为基粒类囊体。
基质类囊体:贯穿在两个或两个以上基粒之间没有发生垛叠的类囊体,称为基质类囊体。
其片层称为基质片层。
◆叶绿体基质:内膜与类囊体之间的区域。
成分:①可溶性蛋白:ribulose-1,5-biphosphate carboxylase/ oxygenase, Rubisco)占类囊体
可溶性蛋白的 80%和叶片蛋白 50%。②DNA③核糖体④脂滴、淀粉粒、植物铁蛋白等
线粒体电子传递:
Mt 中的氧化代谢,是物质最终氧化释能的场所,还原性辅酶彻底氧化。
复习:物质进出线粒体穿梭系统;Malate-aspartate-shuttle;Glycerol-3-phosphotate shuttle;
肉碱与脂肪酸转运;Acetyl-CoA 转运与柠檬酸穿梭
P/O 比值:NADH(FADH2)1 对电子传递到 O2,泵出 10(6)个质子,每合成 1 个 ATP
需 4 个质子
叶绿体光反应、暗反应部位:
光反应部位:在类囊体膜上,由光引起的光化学反应,通过叶绿素等光合色素分子吸收、
传递光能,将水光解生成高能电子,沿电子传递链进行传递,形成 O2 、ATP 和 NADPH
的过程。包括原初反应、电子传递和光合磷酸化。
暗反应部位:在叶绿体基质中,利用光反应产生的 ATP 和 NADPH,使 CO2 还原为糖类
等有机物,即将活跃的化学能最后转换为稳定的化学能,积存于有机物中。这一过程不
直接需要光,故称为暗反应。
半自主性细胞器:自身含有遗传表达系统(自主性);但编码的遗传信息十分有限,其
RNA 转录、蛋白质翻译、自身构建和功能发挥等必须依赖核基因组编码的遗传信息(自
主性有限),故称为半自主性细胞器。
线粒体和叶绿体的半自主性:
很多学者把线粒体和叶绿体的遗传信息系统称为真核细胞的第二遗传信息系统,或核外
基因及其表达体系。这是因为研究发现,线粒体和叶绿体中除有 DNA 外,还有 RNA(mRNA、
tRNA、rRNA)、核糖体、氨基酸活化酶等。说明这两种细胞器都具有独立进行转录和转
译的功能。也就是说,线粒体和叶绿体都具有自身转录 RNA 和翻译蛋白质的体系。但
迄今为止,人们发现叶绿体仅能合成 13 种蛋白质,线粒体能够合成的蛋白质也只有 60
多种,而参与组成线粒体和叶绿体的蛋白质却分别有上千种。这说明,线粒体和叶绿体
中自身编码合成的蛋白质并不多,它们中的绝大多数蛋白质是由核基因编码,在细胞质
核糖体上合成的。也就是说,线粒体和叶绿体的自主程度是有限的,它们对核遗传系统
有很大的依赖性。因此,线粒体和叶绿体的生长和增殖是受核基因组及自身的基因组两
套遗传信息系统控制的,所以它们都被称为半自主性细胞器。
线粒体和叶绿体的起源:
1、内共生起源说:
Mt,cp 起源于原始真核细胞内共生的细菌和蓝藻。
↓
Mereschkowsky,1905 年提出叶绿体起源于细胞内共生的蓝藻;
↓
Margulis,1970 年:线粒体的祖先-原线粒体是一种革兰氏阴性细菌,具有 TCA 循环和电
子传递链;叶绿体的祖先是原核生物的蓝细菌即蓝藻。
线粒体和叶绿体内共生起源说积累较多证据:
(1)基因组在大小、形态和结构方面与细菌相似。不含 5-Methylcytosine,无组蛋白,
自身复制与转录酶
(2)具有独立、完整的蛋白质合成系统,有自己完整的蛋白质合成系统,蛋白质合成
机制有很多类似细菌而不同于真核生物。
(3)mt 和 cp 均以缢裂方式分裂,类似细菌;
(4)两层被膜有不同的进化来源,外膜与细胞的内膜系统相似,内膜与细菌质膜相似,
蛋白质/脂质比接近细菌。
(5)其他佐证,mt 的磷脂成分、呼吸类型和 cyt-c 初级结构均与反硝化副球菌或紫色
非硫光合细菌接近。
(6)发现介于胞内共生蓝藻与叶绿体之间的结构--蓝小体,其特征在很多方面可作为原
始蓝藻向叶绿体演化的佐证。
2、非共生起源说:
真核细胞的前身是一个进化上 比较高等的好氧细菌,通过细胞膜内陷、扩张、分化,
形成 Mt 和 Chl 雏形,经内陷膜包裹形成原始细胞器,进化过程中发生基因组丢失,经
特化为线粒体和叶绿体。
成功之处:解释了真核细胞核被膜的形成与演化的渐进过程
实验证据不多:
(1)无法解释为何线粒体、叶绿体与细菌在 DNA 分子结构和蛋白质合成性能上有那么
多相似之处。
(2)对线粒体和叶绿体的 DNA 酶、RNA 酶和核糖体的来源也很难解释。
第八章 细胞骨架
细胞骨架:细胞骨架是指存在于真核细胞中的蛋白纤维网架体系,包括微丝,微管和中
间丝,均由相应的蛋白亚基组装而成。
狭义:就指胞质骨架,位于细胞质中,由微丝、微管、中间纤维构成纤维型的网络结构。
广义上包括:膜骨架、细胞质骨架、核纤层、核骨架、细胞外基质,构成细胞内外的一
体化网络结构。
微丝的组成、组装:
微丝:又称肌动蛋白纤维, 是指真核细胞中由肌动蛋白组成、直径为 7nm 的骨架纤维。
微丝组成:肌动蛋白(actin)是微丝的结构成分,外观呈哑铃状, 这种 actin 又叫 G-actin,
将 G-actin 形成的微丝又称为 F-actin。
肌动蛋白单体:球状,但中间有一裂缝。裂缝内部有 1 个核苷酸(ATP 或 ADP,但常为
ATP )的结合位点和 1 个二价阳离子(Mg2+或 Ca2+,但常为 Mg2+)的结合位点。
球状肌动蛋白(单体)→纤维状肌动蛋白→微丝
微丝结构:单条直径约为 7 nm 的扭链,由肌动蛋白单体组装而成:在电镜下观察,整
根微丝在外观上是由 2 股纤维以右手螺旋同向盘绕而成,螺距为 36 nm。
组装/去组装条件:
(1)肌动蛋白单体状态:需达到临界浓度;
(2)有成核蛋白 Arp2,Arp3 的参与;
(3)Mg 2+, ATP, Na+, K+较高——G-actin → F-actin,即组装;
(4)Ca2+适当,Na+, K+很低——F-actin → G-actin,即解聚;
极性:正极组装速度大于负极
微管的组成、组装:
电镜-微管 MT,中空管状结构,外径 24nm,内径 15nm.
暂时性结构——间期细胞的细胞质微管;分裂细胞的纺锤体微管等,细胞器和生物大
分子。
永久性结构——纤毛、鞭毛内轴丝微管、轴突内微管束等。
细胞内微管的三种类型:

构成:微管管壁-13 根原纤丝-ab 微管蛋白二聚体纵向排列的异二聚体。
具有极性:组装快的一端正极,慢的一端负极。
微管的组装和解聚:
装配三个时期:
(1)成核期:由α、β微管蛋白聚合成寡聚体核心,接着二聚体在其两端和侧面增加
使之扩展成片状带,加宽成 13 根原纤维即构成一段微管。
(2)聚合期:又称为延长期,该期微管蛋白聚合速度大于解聚速度,微管延长。
(3)稳定期:微管的聚合和解聚速度相等。
踏车行为:指在微丝组装/去组装过程中,正极由于肌动蛋白亚基的不断添加而延长,
负极由于肌动蛋白亚基的去组装而缩短,总长度几乎不变的现象。
微管组织中心:在活细胞内,能够起始微管的成核作用,并使之延伸的细胞结构即谓之
microtubule organizing center, MTOC。如中心体、轴突、基体和其它特殊的部位(核膜外
表面、细胞的两极、高尔基体的反面膜囊区和新断的微管)等。
马达蛋白:即能够利用水解 ATP 释放的能量驱动自身有规则地沿微丝或微管定向运动的
蛋白。
马达蛋白类型:沿微丝运动的肌球蛋白(myosin)、沿微管运动的驱动蛋白(kinesin)
和动力蛋白(dynein)。
结构特征:马达蛋白具有 2 种结构域:①与微丝或微管结合的马达结构域;②与大分子
复合物或膜性细胞器特异结合的“货物”结构域。
驱动蛋白:指能利用 ATP 水解所释放的能量驱动自身及所携带的“货物” (如膜性细
胞器)沿微管运动的一类马达蛋白,与细胞内物质运输相关。
结构特征:驱动蛋白分子是一条长 80nm 的杆状结构,由头部(即重链的 N 端,具有 2
个球状的马达结构域)、颈部(重链)、杆状区(重链)和扇形尾端(即重链的 C 端和
轻链构成,具有“货物” 结合结构域)组成。
微管结合蛋白:一类结合在微管表面的蛋白质,始终伴随微管的组装和去组装而存在,
对微管的组织结构和功能具有调控作用。包括 MAP1、MAP2、MAP3、MAP4 和 tau 蛋白。
细胞松弛素、鬼笔环肽对微丝的作用:
细胞松弛素对微丝的作用:可切断微丝,并和微丝末端结合,抑制 actin 聚合,不影响
解聚,抑制运动
鬼笔环肽对微丝的作用:只与 F-肌动蛋白结合,抑制微丝解聚,使其保持稳定状态-标
记微丝!
秋水仙素、紫杉醇等对微管的作用:
秋水仙素对微管的作用:低浓度的秋水仙素处理细胞,可立即破坏细胞内的微管或纺锤
体的结构。秋水仙素在微管末端的结合影响该处的组装,但并不影响该处的去组装。
紫杉醇对微管的作用:作用与秋水仙素相反,即不影响微管的组装,但阻止微管的去组
装。
中间丝组分、组装:
中间丝组分:I 型角蛋白:位于胞内, 酸性,亦称胞质角蛋白
II 型角蛋白:中/碱性,头发、指甲等坚韧结构所有。二者以异源二聚体形式参与上皮细
胞中间丝组装。
III 型中间丝:波形纤维蛋白(间充质细胞及中胚层来源的细胞),一端与核膜相连,另
一端与细胞表面的桥粒、半桥粒相连,将细胞核和细胞器维持在特定空间、结蛋白等。
IV 型:3 种神经丝蛋白亚基
V 型:核纤层蛋白 A 等
IV 型:巢蛋白、微管卷曲蛋白等。
中间丝组装:

第九章 细胞核与染
色质
核被膜:(nuclear envelope)位于细胞核最外层,是细胞核与细胞质之间的界膜。
功能:(1)屏障,核质两大功能区的界限(2)细胞核内外物质、信息交换与交流的调
控
构成:双层核膜,核孔复合体,核纤层
核被膜在细胞周期中的崩解与组装:
分裂期:双层核被膜崩解为单层膜泡,核孔复合体解体,核纤层去组装
分裂末期:核被膜围绕染色体重新形成
处于动态变化之中,新核膜来自旧核膜的重建
核孔复合体:核孔上镶嵌的复杂结构谓之核孔复合体,数目呈现动态变化。
NPC 组成成分
总分子量大小:1.25×108,30 多种多肽,1000 多个蛋白质分子。
核孔蛋白(nucleoporin, Nup):指所有的核孔复合体蛋白,进化上高度保守。 P181 TAB
生化与遗传分析可以鉴定,酵母是良好材料。
1、gp210:第一个被鉴定,结构性跨膜糖蛋白,位于孔膜区,210KD,在糙面内质网上
合成,含 ConA 结合的 N-甘露糖残基寡糖链,锚定核孔复合体。
功能:
(1)介导核孔复合体与核被膜的连接,将核孔复合体锚定在“孔膜区”,从而为核孔复
合体装配提供一 个起始位点
(2)在内、外核膜融合形成核孔中起重要作用
(3)在核孔复合体的核质交换功能活动中起一定作用。抗 gp210 核周间隙内肽段抗体
可降低蛋白质入核转运速度。
是跨膜运输复合体,是核质交换的双向性亲水通道。
运输特点:
(1)双功能: 主动运输、被动运输
(2)双向性:蛋白质入核;RNA,RNP,核糖体亚单位出核运输
运输方式:(1)被动运输,<50KD,自由扩散,4℃不被抑制,无需能量。大小无限制,
协助扩散, 4℃被抑制,与 NPC 相互作用
(2)主动运输,信号介导核输入,核定位信号 NLS 及其受体/载体, 4℃被抑制。核输
出,核输出信号 NES 及其受体/载体, 4℃被抑制。
亲核蛋白:是指在细胞质中合成,需要或者进入细胞核内发挥功能的一类蛋白质. 核定位序列:(NLS)指亲核蛋白所含的一段特殊氨基酸序列,富含碱性氨基酸残基,可为
亲核蛋白向核内转运提供定向和定位作用。
NLS 特点:
NLS 的氨基酸残基片段可以是一段连续的序列(T 抗原),也可以分成两段,两段之
间间隔约 10 个氨基酸残基(核质蛋白)。
NLS 序列可存在于亲核蛋白的不同部位,在指导完成核输入后并不被切除(与信号肽
区分)。
NLS 只是亲核蛋白入核的一个必要条件而非充分条件
核输出信号:一些与 RNA 共同出核的蛋白因子,含有对他们的出核转运起决定作用的
氨基酸序列。
核小体:核小体结构要点 p189
(1)每个核小体单位包括 200bp 左右的 DNA 超螺旋和一个组蛋白八聚体及一个分子
H1
(2)组蛋白八聚体构成核小体的盘状核心结构
(3)146bp 的 DNA 分子超螺旋盘绕组蛋白八聚体 1.75 圈, 组蛋白 H1 在核心颗粒外结
合额外 20bp DNA,锁住核小体 DNA 的进出端,起稳定核小体的作用。
(4)两个相邻核小体之间以连接 DNA 相连,典型长度 60bp,不同物种变化值为 0~80bp
(5)组蛋白与 DNA 之间的相互作用主要是结构性的,基本不依赖于核苷酸的特异序列,
(6)核小体沿 DNA 的定位受不同因素的影响,
染色质:是指间期细胞核内由 DNA、组蛋白、非组蛋白及少量 RNA 组成的线性复合结
构,能被碱性染料强烈着色,是间期细胞遗传物质存在的形式
染色体:是指细胞在有丝分裂或减数分裂的特定阶段,由染色质聚缩而成的棒状结构。
染色体的基本结构元件:自主复制 DNA 序列,着丝粒 DNA 序列,端粒 DNA 序列
常染色质和异染色质及其区别:
(1)常染色质:指间期核内染色质纤维折叠压缩程度低, 处于伸展状态, 用碱性染料染
色时着色浅的那些染色质. (2)异染色质:指间期细胞核中,用碱性染料着色深,折叠压缩程度高,处于聚缩状
态的染色质组分。
区别:构成:常染色质由单一序列 DNA 和中度重复序列 DNA。异染色质由相对简单、高
度重复的 DNA 序列构成。
活性:常染色质并非所有基因都具有转录活性,常染色质状态只是基因转录的必要条件
而非充分条件。异染色质具有显著的遗传惰性, 不转录也不编码蛋白质。
复制:异染色质与常染色质相比表现为晚复制早聚缩。
非组蛋白:指与特异序列 DNA 相结合的蛋白质,又名序列特异性 DNA 结合蛋白。
特性:(1)多样性(2)识别 DNA 特异性,即序列结合特异性;(3)功能多样性
组蛋白:真核生物染色体的基本结构蛋白,富含带正电荷的 Arg 和 Lys 等碱性氨基酸,
属碱性蛋白质,可以和酸性的 DNA 紧密结合
活性染色质、非活性染色质及其结构和表达特征:
活性染色质:具有转录活性的染色质,结构疏松,便于转录因子和聚合酶与 DNA 结合
和移动。由于结构疏松,H3 暴露出巯基,可与有机汞结合进行亲和层析,再用 DTT 洗
脱
非活性染色质:是没有转录活性的染色质。
活性染色质特征:1、活性染色质对 DNase I 超敏感。2.DNase I 消化可将染色质降解为酸
溶性 DNA 小片段
活性染色质蛋白质组成与修饰变化
(1)生化分析
活性染色质很少有组蛋白 H1 与其结合;
活性染色质的组蛋白乙酰化程度高;
活性染色质核小体组蛋白 H2B 很少被磷酸化;
活性染色质中核小体组蛋白 H2A 在许多物种很少有变异形式;
组蛋白 H3 变种 H3.3 只在活跃转录的染色质中出现
HMG14 和 HMG17 只存在于活性染色质中,与 DNA 结合。
DNase l 超敏感位点:又称 DNA 酶|超敏感位点,在对染色质进行低 DNasel 处理,切割
将先发生在少数特异性位点.上,这些特异性位点叫做 DNasel 超敏感位点。
表观遗传学:指在基因的 DNA 序列没有发生改变的情况下,基因功能发生了可遗传的
变化,并最终导致了表型的变化。
核型:是指染色体组在有丝分裂中期的表型, 包括染色体数目、大小、形态特征的总和。
核仁周期:核仁的形态、大小、数目随细胞周期不同而变化的现象。
核仁在细胞周期中的动态变化:进入有丝分裂时,核仁首先变形、变小,随染色体凝集,
核仁消失,rRNA 合成停止,中期及后期没有核仁;末期 rRNA 合成重新开始,核仁物质
凝聚成分散的前核仁体,在 NOR 周围融合形成正在发育的核仁。其动态变化依赖于正
在转录的 rRNA 基因和 RNA Pol I 的活性形式. 核纤层:核纤层普遍存在于高等真核细胞中,是内层核被膜下纤维蛋白片层,其纤维直
径为 10 毫微米左右,纤维纵横排列整齐呈纤维网络状。
第十章 核糖体
多核糖体:具有特姝功能与形态结构的核糖体与 mRNA 的聚合体称为多核糖体。
第十一章 细胞信号传导
细胞通信:
概念:细胞产生的胞外信号与靶细胞表面相应的受体结合,引发受体构象变化,进而导
致细胞内信号通路建立,最终调节靶细胞功能。
类型:(1) 分泌化学信号:长距离或短距离;内分泌;旁分泌;自分泌;化学突触传
递神经信号;外激素至其它个体细胞。
(2)细胞间接触依赖型通讯:细胞间直接接触,质膜上信号分子与靶细胞质膜受体分
子相互作用,信号分子与受体都是细胞的跨膜蛋白。细胞-细胞黏着、细胞-基质黏着。
发育中相邻细胞分化。
(3)动物细胞间的间隙连接和植物细胞间的胞间连丝,小分子交换的代谢耦联
信号分子:
1、定义:信号分子(signal molecule):细胞的信息载体,包括化学信号(激素、局
部介质、神经递质等)、物理信号(声、光、电、温度)。
激素:激素是内分泌细胞合成的化学信号分子,它们经血液循环被送到体内各个部位作
用于靶细胞。
神经递质:突触前端释放的一种化学物质,与突触后靶细胞结合,并改变靶细胞的膜电
位。
局部介质:即由各种不同类型的细胞合成并分泌到细胞外液中的信号分子,它只作用于
周围的细胞,如生长因子、淋巴因子和前列腺素等。
化学信号种类按化学性质分类:
(1)气体性信号分子(可自由扩散进入细胞直接激活效应酶,如 NO);
(2)疏水性信号分子-可穿过细胞质膜与细胞内受体结合成激素-受体复合物来调节基
因表达,如甾类激素和甲状腺素;
(3)亲水性信号分子-不能穿过质膜,只能通过与靶细胞表面受体结合而启动细胞信号
转导,如大多数蛋白类激素、神经递质和局部介质。
(4)膜结合信号分子:表达在细胞质膜上,与靶细胞质膜上受体分子相互作用,产生
应答反应
信号转导:
信号转导是实现细胞间通讯的关键过程。
(细胞)信号转导(signal transduction):指细胞将外部信号转变为自身应答反应
的过程。 这是实现细胞间通讯的关键过程。
受体:
1、受体:能够识别和选择性结合某种配体的大分子,绝大多数为糖蛋白,少数是糖脂
(霍乱毒素等)或糖蛋白糖脂复合物(TSH 受体)
2、分类:
(1)按部位分类:
细胞内受体: 存在于胞质或核基质中,为胞外亲脂性信号分子所激活:类固醇、甲状腺
素,视黄酸;
细胞表面受体: 位于质膜表面,胞外亲水性信号分子所激活:分泌型信号分子受体、膜
结合型信号分子。
(2)按信号传导机制,表面受体三大家族
①离子通道欧联受体:本身既有配体结合位点,又是离子通道,无需中间步骤即可实现
跨膜信号转导,配体门离子通道。
②G 蛋白偶联受体:GPCR,种类多
③酶联受体:受体胞内结构域具酶活性,或受体包内段与酶相联系。
受体结构特征:在结构上至少具备两个功能域:配体结合功能域、效应功能域
3、受体作用特点:
(1)高度的亲和力(high affinity):激素及细胞因子等配体与其受体的结合具有高度
亲和力。通常用其解离常数(Kd)来表示其亲和力的大小,多数配体与受体的解离常数
为 10-11~10
-9 mol/L。
(2)高度的特异性(high specificity):指一种激素或细胞因子只能选择性与相应的
受体结合的性质。
(3)可逆性(reversibility):配体与受体通常通过非共价键而结合,因此可以采用简
单的方法将二者分离开。
(4)可饱和性(saturability):由于存在于细胞膜上或细胞内的受体数目是一定的,
因此配体与受体的结合也是可以饱和的。
(5)特定的作用模式:在不同细胞中,受体的种类和含量分布均不同,表现为特定的
作用模式。不同细胞可以不同方式应答同一化学信号,Ach 作用于骨骼肌、心肌和唾液
腺效应不同;血糖调节一种细胞有一套多种类型的受体,应答多种信号,启动不同效应.
(6)受体的脱敏性:可因受体本身或受体下游蛋白的变化,从而导致机体适应性。
第二信使:
1、第二信使定义:胞内产生的非蛋白类小分子,其浓度变化应答胞外信号与细胞表面
受体的结合,调节细胞内酶和非酶蛋白的活性,从而在细胞信号传导过程中行使信号放
大功能,调控细胞活动。
2、第二信使种类:环核苷酸类(cAMP, cGMP),Ca2+,脂类衍生物(DAG(二酰基甘油),
花生四烯酸,神经酰胺等);糖类衍生物(IP3(1,4,5-肌醇三磷酸)。
分子开关:
(1)定义:分子开关是指通过活化(开启) 和失活(关闭)两种状态的转换来控制下
游靶蛋白活性的一类调控蛋白。在进化上非常保守。
(2)分子开关类型:
①GTPase 开关蛋白超家族(Gilman & Rodbell,94 年炸药奖),鸟苷酸结合蛋白,包
括三聚体 GTP 结合蛋白和单体的 Ras( rat sarcoma )。通过与 GTP 或 GDP 结合,实现
开或者关的状态,从而调控靶蛋白活性。
需要 GEF(鸟苷酸交换因子)的介导。
②蛋白激酶与蛋白磷酸酶(Krebs & Fischer 1992 诺奖),使靶蛋白磷酸化去磷酸化。
③钙调蛋白(CaM)与 Ca
2+的结合或解离产生的激活或失活状态,具有结合钙离子并介导
其许多细胞内信号途径的细胞内小分子蛋白,每个CaM分子具有4个Ca2+离子结合位点,
CaM 通过与 Ca2+的结合或解离而分别处于活化或失活的“开启”或“关闭”状态,Ca2+-CaM
复合物可结合多种酶及其它靶蛋白, 修饰其活性。
信号转导系统的主要特性:
(1)特异性(specificity):包括“结合” 特异性、饱和性和可逆性以及“效应器”
特异性。
(2)放大效应(amplification):效应器蛋白(酶或离子通道蛋白)引发级联反应(放
大效应)。
(3)网络化效应与正反馈(positive feedback)和负反馈(negative feedback)环
路:网络化效应有利于克服分子间相互作用的随机性对细胞生命活动的负面干扰;正反
馈和负反馈环路对于及时校正反应的速率和强度是最基本的调控机制。
(4)整合作用(integration):细胞通过整合不同的信息,对细胞外信号分子的特异
性组合做出程序性反应,甚至做出生死抉择,这样才能维持生命活动的有序性。
G 蛋白耦联受体:
一、G 蛋白耦联受体的结构与激活
GPCR 是细胞表面最大的受体家族,指配体-受体复合物与靶蛋白的作用要通过与 G 蛋白
的耦联,在细胞内产生第二信使,从而将胞外信号 k 跨膜传递到胞内,产生特定生理效
应。嗅觉、视觉、血糖、动作电位等均依赖 GPCR,30%处方药要针对 GPCR 研发。
G 蛋白:三聚体 GTP 结合调节蛋白的简称。
1、G 蛋白组分:3 聚体 GTP 结合蛋白,Ga,Gb,Gg 三聚体,铆定于质膜上,Ga 是分子
开关蛋白,具 GTPase 活性。配体与受体结合,三聚体解聚,Ga-GTP 活化开启,结合并
激活效应器蛋白。Ga-GTP 水解形成 Ga-GDP 时,失活,3 聚体再组装。(1994 年诺奖 Gilman
& Rodbell)
2、类型:G 蛋白主要是两类,即激活型 G 蛋白(Gs)和抑制型 G 蛋白(Gi)。G 蛋白(Gs
或 Gi)的效应酶主要是腺苷酸环化酶(AC),可催化第二信使 cAMP 的产生,从而将胞
外的信号传递到细胞内。
G 蛋白耦联受体介导的细胞信号通路:
按效应器蛋白,GPCR 分为三类:
(1)激活离子通道的 G 蛋白偶联受体所介导的细胞信号通路
(2)激活或抑制腺苷酸环化酶的 G 蛋白偶联受体所介导的细胞信号通路
(3)激活磷脂酶 C(以 IP3 和 DAG 作为双信使)的 G 蛋白偶联受体所介导的信号通路
许多离子受体是 G 蛋白耦联受体,其效应器是钠离子或钾离子通道,从而改变膜电位。
(一)激活离子通道的 G 蛋白偶联受体所介导的细胞信号通路
1. 心肌细胞上 M 型(蝇蕈碱)Ach 受体激活 G 蛋白开启 K+通道-抑制性 G 蛋白
Ach→M 型 Ach 受体→Gi 蛋白(GiαGβγ)→ Gβγ结合 →K+通道打开→ K+外流→超
极化→减缓心肌细胞的收缩频率
横纹肌的 N-型 Ach 受体—钠离子通道打开—钠离子内流-去极化。
G 蛋白不同!!!
2. 激活 Gt 蛋白偶联的光敏感受体诱发 cGMP-门控阳离子通道-视觉的形成
Gt=G 蛋白偶联的视紫红质(transducing, 传导素)
光→视紫红质(感受弱光刺激)→ Gt 蛋白(传导素)(GtαGtβγ)→ Gtα→ cGMP-PDE
(磷酸二酯酶)抑制性γ亚基→ cGMP-PDE(α/β) →破坏 cGMP(水解)→ cGMP 门
控阳离子通道关闭→膜瞬间超极化→视神经→脑
(二)激活或抑制 AC 的 G 蛋白偶联受体
普遍机制:激活 AC,调控 cAMP,下游调控
Gα的首要效应酶是 cAMP 环化酶(AC),调控 cAMP 水平,作用于下游分子。
可以是激活或者抑制 cAMP 环化酶,涉及 5 个蛋白组份:
(1)刺激性激素受体(Rs)(2)抑制性激素受体(Ri)(3)刺激性 G 蛋白(Gs)(4)
抑制性 G 蛋白(Gi)(5)腺苷酸环化酶(AC)
系统组成
1、腺苷酸环化酶:跨膜 12 次。在 Mg2+或 Mn2+的存在下,催化 ATP 生成 cAMP;环腺苷
酸磷酸二酯酶 cAMP phosphodiesterase, PDE):降解 cAMP 生成 5’-AMP,起终止信号
的作用。
2、PKA
cAMP 升高激活 PKA,PKA 可促使多种酶或蛋白质丝氨酸或苏氨酸残基的磷酸化,从而调
节酶的催化活性或蛋白质的生理功能。PKA 由四个亚基构成(R2C2),两个催化亚基(C),
两个调节亚基(R)。当 R 与 cAMP 结合后发生变构(每一调节亚基可结合两分子 cAMP),
与催化亚基解聚,从而激活催化亚基。
G 蛋白耦联的 cAMP-PKA 信号转导过程:激素→G-蛋白偶联受体→G-蛋→AC↑→cAMP ↑
→cAMP 依赖的蛋白激酶 A→基因调控蛋白→基因转录
cAMP-PKA 对肝细胞和肌细胞糖原代谢的调节(快速的调控过程)
(三)激活磷脂酶 C(PLC)(以 IP3 和 DAG 作为双信使)的 G 蛋白偶联受体介导的信号
通路
(1)磷脂酰肌醇代谢途径:双信使 DAG 和 IP3 来自膜结合的磷脂酰肌醇(PI)。磷脂
酰肌醇(PI)(PI-4K 催化)→磷脂酰肌醇-4-磷酸(PIP)(PI-5K) →磷脂酰肌醇-4,5-
二磷酸(PIP2)(PLC) →二酰甘油(DAG)+ 肌醇-1,4,5-三磷酸(IP3) → DAG 是亲
脂性分子,锚定在膜上; IP3 扩散至细胞质中。催化从 PIP2 到 DAG 和 IP3 是由 PLC 水
解完成
(2)磷脂酶 C(PLC)的β异构体(PLC β):膜镶嵌蛋白,依赖 Ga 激活,催化 PIP2
水解产生二酰甘油(DAG)和肌醇-1,4,5-三磷酸(IP3)2 个第二信使分子。
(3)蛋白激酶 C(PKC):为 Ca2+激活型蛋白激酶,属于丝氨酸/苏氨酸蛋白激酶家族,
可广泛磷酸化多种不同的蛋白质底物。
1、IP3-Ca2+信号通路
胞外信号分子结合 CPCR, Goα或 Gqα被激活,然后去激活 PLC,催化 PIP2 水解生成
IP3 和 DAG 两个第二信使,故为双信使通路。IP3 扩散并与 ER 上 IP3 敏感的钙离子通道
蛋白结合,钙离子释放进入细胞质基质。在这一途径中, 细胞质基质 Ca2+浓度升高又
促进钙离子通道开放,Ca2+浓度呈现爆发性的增高;形成正反馈,钙离子泵又把钙离子
泵回 ER 腔,因而产生 Ca2+振荡。
钙火花(calcium spark)与 IP3- Ca2+信号通路关系
(1)IP3 调控 ER Ca2+的专一性:钙离子通道与 IP3 结合,导致通道开放;借助 confocal
与 Fura-2,Fluo-3,可记录钙离子传递机制。
(2)钙火花: Ca2+ 荧光探针 Fluo-3 荧光强度在局区域约 10ms 骤升一倍后,又会在
20ms 消失的现象。
(3)效应:单个心肌细胞每次收缩,形成 104 个钙火花; Ca2+通过与 CaM 结合,发挥
调控作用。
3、DAG-PKC 信号通路
DAG-PKC 依赖于 Ca2+和磷光素酶。
DAG-PKC 通路→激活 PKC→使靶蛋白上的丝氨酸和苏氨酸磷酸化。(MAP3K,NF-kB)-- 调节转录
PKC:存在于胞液中,可催化底物蛋白质丝氨酸或苏氨酸残基的磷酸化,有 12 种同工酶。
经典的蛋白激酶 C 需在 Ca2+,DAG 和磷脂酰丝氨酸(PS)的存在下才能被激活。
酶联受体:又称催化性受体,当胞外配体与受体的胞外部分结合时即激活受体胞内结构
域的酶活性,包括两大类:RTK 和细胞因子受体。
种类:这类受体细分包括 5 类:
1 受体酪氨酸激酶;② 受体丝氨酸/苏氨酸激酶;③ 受体酪氨酸磷酸酯酶;④受体鸟
苷酸环化酶;⑤ 酪氨酸蛋白激酶联的受体。
粘着斑 (Focaladhesion):细胞通过肌动蛋白纤维与借助整联蛋白与细胞外基质之间的
连接方式
整联蛋白:普遍存在于脊椎动物细胞表面、异亲型结合,依赖 Ca+或 Mg2+的细胞黏着蛋
白, a 和β两个亚基形成跨膜异源二聚体,为钙离子或镁离子依赖性黏着分子,介导细
胞与细胞之间或细胞与基质之间的黏着。人体细胞中已发现 24 种α链和 9 种β链,它
们相互配合形成多种二聚体整联蛋白,可与不同的配基结合,从而介导细胞与基质、细
胞与细胞之间的粘着。作用机制:亚基在胞内与肌动蛋白相互作用,胞外部分具有
RGD(Arg-Gly-Asp)结构域,与胞外基质结合整联蛋白介导细胞与基质之间黏着的典型结
构有黏着斑和半桥粒。
功能:(1)细胞黏着 (2)作为受体介导信号从胞外向胞内传递,即依赖于细胞内
tyrosine kinase-FAK 的信号传导作用。 (3)促进细胞增殖。培养细胞依赖整联蛋白
贴附于胞外基质上才能正常生长,否则就会停止分裂直至凋亡,即锚定依赖性生长。
第十二章 细胞周期与细胞分裂
细胞增殖(cell proliferation):是细胞生命活动的重要特征之一,通过细胞分裂,使得
细胞数目增加。
意义:
(1)是生物繁育和生长发育的基础。单细胞生物,增殖增加个体数量,多细胞生物,
增殖与分裂分化密切相关。
(2)成体生物细胞通过增殖,弥补代谢细胞损失;
(3)替代衰老死亡细胞,维持机体细胞数量平衡;
(4)创伤愈合、组织再生、病理组织修复,依赖细胞增殖;
(5)细胞增殖受到
严密调控。
细胞周期:从细胞一次有丝分裂结束开始,经过物质积累,到下一次有丝分裂完成所经
历的一个有序过程,称作一个细胞周期。
标准细胞周期的各时相组成、事件、特点:
1、G1 期:第一阶段,从上一次分裂形成子代细胞开始到 DNA 合成。细胞生长,合成
蛋白质、糖类、脂质等。
G1 晚期存在着一个 checkpoint(检查点),若通过,即进入 S 期,进行 DNA 合成。
影响通过 G1 checkpoint 因素:
外在:营养、激素刺激
内在:细胞分裂基因(cell division cycle gene, cdc)表达调控。
细胞周期检查点 Checkpoint:在细胞周期中,对细胞周期进行特异性监控的关键时期。
包括 G1/S,S,G2/M 和纺锤体检查点。意义:保持基因组稳定性
2、S 期:DNA 合成,组蛋白往往同步进行合成
3、G2 期:染色体倍性由 2n 变为 4n,需要合成一些蛋白质和 RNA 分子。
G2/M 期检查点:检查 DNA 是否完成复制,损伤是否修复,细胞大小和营养物质积累
及其环境因素等。若均通过,即进入 M 期。
4、M 期:细胞分裂,包括 mitosis and meiosis. 细胞周期的运转:CDK 调控系统
5、细胞周期长短测定
细胞周期长短随细胞种类环境条件等而变化。
(1)标记有丝分裂百分比法(Percentage labeled mitosis, PLM)
使用 3H-TdR 短期培养细胞,将 3H-TdR 洗脱,每隔 0.5-1h,定时取样,放射自显影观
察,测定带放射性 M 期标记细胞的百分比。
细胞分裂指数:细胞群体中分裂细胞所占的百分比,表示细胞增殖旺盛程度的指标。
凡是处于 S 期均被标记,这些细胞会进入 M 期,S 后期,早进入 M,S 前期,晚进入
M。
TG2 期:从加入 3H-TdR 时 PLM=0,到第一个放射性 M 期开始出现所经历的时间,即
最晚期 S 期进入 M,时间正好为 G2 期。
TM 期:S 期逐渐进入 M,PLM 逐渐达到 100%,从出现被标记 M 期到 M 期达到 100%,
所需要的时间。
总周期 Tc:从开始出现放射性 M 期到下一次出现放射性 M 期的时间
TG2-G2 期时间,TM-M 期时间,TS-S 期时间,TC-细胞周期时间
Ts:PLM=100%会维持一段时间,当 PLM 开始下降时,说明 S 早期标记的细胞也进入 M
期,从出现 M 期,到 PLM 开始下降的时间,即为 Ts
Tc:从第一次出现 PLM 到下一次出现 PLM 之间的间隔,即为一个细胞周期时间。
TG1= TC -(TG2 + TM + TS )
但由于细胞周期不同步,第一个 PLM 峰达不到 100%,以后的峰会衰减,PLM 也不定会
下降到 0,故常用(TG2 + 1/2TM) - TM 求 TM。
(2)流式细胞仪测定方法
根据 DNA 含量分类细胞,根据类群细胞的数量推断时间。即根据 DNA 在不同时间内的
变化,确定细胞周期时间长短。G1, G2/M 期分别为 1C 和 2C(2n & 4n),S 期介于 1C 和
2C 之间。
细胞周期中的细胞群体:
(1)周期中细胞:可以持续分裂、细胞周期持续运转的细胞,上皮组织的基底层细胞,
干细胞;
(2)静止期细胞:停止分裂执行功能,通常在 G1 期转化为 G0 期细胞,结缔组织中成
纤维细胞,一旦接受信号刺激,就可进入细胞周期
(3)终末分化细胞:失去分裂性能,RBC,神经细胞
细胞周期同步化方法:
(一)自然同步化
(二)人工同步化—人工选择或人工诱导
人工选择同步化:是指人为地将处于不同时相的细胞分离开来,从而获得不同时相的细
胞群体。
1、有丝分裂选择法:单层培养细胞,当处于对数期生长时,处于分裂期(M 期)细胞变圆,
贴壁能力变弱,轻轻振荡培养瓶,M 期细胞即从瓶壁脱落,悬浮于培养液中,收集培养
液,离心后即获得同步化 M 期细胞。优点:操作简单,同步化程度高,效率高,细胞
未经药物处理,不受药物伤害。缺点:获得的细胞数量较少。
2、DNA 合成阻断法:使用对细胞周期其它时相无影响的 DNA 合成抑制剂,特异性抑制
DNA 合成,最终可将细胞群阻断在 S 期或 G1/S 交界处。其中过量 TdR 对 S 期细胞的毒
性较小,因此常用 TdR 双阻断法诱导细胞同步化。通过第二次抑制,周期阻断在 G1/S
期。
优点:同步化程度高,适用于任何培养体系,用途广,可将几乎所有的细胞同步化。
缺点:产生非均衡生长,个别细胞体积增大。需对该细胞周期各时相了解清楚
3、分裂中期阻断法:利用破坏微管的药物将细胞阻断在中期,常用的药物有秋水仙素
和秋水仙酰胺,后者毒性较少。在药物持续存在条件下,分裂期细胞数量逐渐累加,经
离心分离,获得中期细胞。优点:操作简便、效率高且无非均衡生长现象。缺点:毒性
相对较大,可逆性较差,处理时间过长,则不能恢复细胞周期运转。
(4)条件依赖性突变株:将与细胞周期调控有关的条件依赖性突变株转移到限定条件
下培养,所有细胞便被同步化在细胞周期中某一特定时期。
有丝分裂各时相:
1、前期事件:
(1)染色质凝缩,两条染色单体在着丝粒处相连;细胞骨架解聚,中心体周围微管大
量组装;高尔基体、内质网等解体。在前期染色体凝缩阶段,需要保持已经复制的两条
姐妹染色单体的凝缩和黏连。凝缩蛋白和黏连蛋白,是 SMC(structural maintenance of
chromosome)家族具 ATPase 功能的蛋白,在这一过程具有重要作用。
(2)细胞分裂极的确定和纺锤体装配:分裂极的确定与中心体复制、分离密切相关。
间期动物细胞含一个 MTOC,即中心体,在 S 期末,两个中心粒在各自垂直的方向复制
出一个中心粒,形成两个中心体。当前期开始时,2 个中心体移向细胞两极,并同时组
织微管生长,由两极形成的微管通过微管结合蛋白在正极末端相连,最后形成有丝分裂
纺锤体。
2、前中期
(1)标志事件之一:核膜崩解,形成小泡,核纤层解聚为核纤层蛋白,染色体进一步
浓缩,变粗变短,朝纺锤体赤道面移动。核纤层蛋白磷酸化促进了崩解,与 MPF 密切
相关,MPF 促进 Ser22,192 磷酸化;Lamin A 以溶解状态存在,B 与核膜小泡结合。
(2)标志事件之二:完成纺锤体装配,形成有丝分裂器。
动粒成熟,星体微管正极捕获染色体,与染色体一侧的动粒结合,形成动粒微管,纺锤
体形成,染色体向赤道面运动。
动物细胞有丝分裂器:由星体微管,染色体动粒微管和极微管及其结合蛋白构成。
因动粒微管长度不等,染色体尚未排列在赤道面上。
纺锤体微管组装十分复杂 微管在中心体周围组装及已完成复制的中心体分离; 分
离过程需要驱动蛋白相关蛋白(KRPs)和胞质动力蛋白的作用,前者负向运动,后者向
微管正极移动(看马达结构域的结合位点)。
(3)标志事件之三是染色体整列
染色体整列:前中期染色体在动粒微管的牵拉下,向赤道面上运动的过程。这一过程需
要 Mad2 和 Bub1 蛋白参与。
动粒外侧为纺锤体微管提供附着,内侧与着丝粒交错。故每条中期染色体上有两个动粒,
将被平均分到两个子细胞中。
3、中期
主要标志事件:染色体整列完成,所有染色体均排列在赤道面上。
染色体整列两个学说:牵拉假说,外推假说
中期染色体动粒分别面向两极;每个动粒上结合的纺锤体微管可达到几十支
4、后期
两条染色单体相互分离,形成子染色体,并分别向两极运动。动粒微管变短,极性微管
加长,两极之间距离拉长。1-2m/min
后期的两个阶段:A 段、B 段
后期 A:动粒微管变短,牵拉姐妹染色单体向两极移动。
机制:靠动粒端微管解聚,动力蛋白沿微管向极部运动。
后期 B:极微管变长,两极之间距离拉长。
机制:驱动蛋白相关蛋白(KRPs)结合极微管,并向微管正极移动,使得来自两极的极
微管重叠区之间滑动。
染色体运动速度:1-2m/min
5、 末期:
染色单体到达两极,动粒微管消失,极性微管继续加长。染色单体去浓缩,核膜围绕每
个染色单体重新组装,经膜泡融合形成子核,同时,NPC 组装,核仁重新组装,RNA 合
成恢复。 Golgi 体和 ER 重新形成并生长。
6、 胞质分裂:cytokinesis
始于分裂后期,完成于细胞分裂末期。动物细胞赤道板细胞表面下陷,形成分裂沟
(furrow),逐渐加深直至两个子细胞分开。分裂沟位置与纺锤体位置相关。
(1)肌动蛋白、肌球蛋白参与分裂沟的形成和胞质分裂,在分裂沟下方,由肌动蛋白、
微管、小膜泡等聚集,共同构成一个环形致密层,谓之中间体(midbody)。
(2)收缩环:胞质分裂开始时,大量肌动蛋白和肌球蛋白在中间体处组装成微丝并相
互组成微丝束,环绕细胞,谓之 contractile ring. (3)动物细胞胞质分裂 4 个步骤 分裂沟位置的确立; 肌动蛋白聚集和分裂环形成;
收缩环收缩; 收缩环处细胞膜融合并形成两个子细胞
中心体列队(整列):中心体分离时,负向运动的马达蛋白在来自姐妹中心体微管之间
搭桥,通过向负极运动,将被结合的微管牵拉在一起,组成纺锤体微管,中心体位于两
极
动粒:每条中期染色体上 2 个,位于着丝粒两侧,S 期,动粒复制。动粒为染色体向两
极运动所必需
纺锤体微管:星体微管、动粒微管、极微管
减数分裂:有性生殖细胞分裂时,染色体只复制一次,而细胞连续分裂两次,导致染色
体数目减半的特殊分裂方式。
二价体:两条配对同源染色体紧密结合在一起形成的结构。
联会:同源染色体配对的过程。联会可以同时发生在同源染色体的几个点上
联会复合体:指同源染色体联会部位形成的特殊复合结构,与同源染色体联会和基因重
组有关。
端化:终变期 diakinesis——染色体重新凝集,形成短棒状结构,交叉向染色体臂的端部
移行,其过程谓之端化,到终变期末,同源染色体仅在其端部和着丝粒处相互连结。
动物细胞胞质分裂四个步骤:分裂沟位置的确定,肌动蛋白聚集和收缩环形成,收缩环
收缩,收缩环处细胞膜融合并形成两个子细胞
第十三章 细胞增殖调控与癌细胞
MPF 的发现及其作用:
MPF (maturation-promoting factor, mitosis-promoting factor, M phrase-promoting factor)。
1970,Johnson and Rao Hela 细胞融合,M 期与间期细胞融合,导致间期染色体凝集;
即早熟染色体凝集 PCC。
PCC:即早熟染色体凝缩,主要是指与 M 期细胞融合的间期细胞(G1、S 和 G2)发生的
形态各异的染色体凝缩。G1 期 PCC 为细单线状(因 DNA 未复制),S 期 PCC 为粉末状
(因 DNA 由多个部位开始复制),G2 期 PCC 为双线染色体状(说明 DNA 复制已完成),
这样的形态变化可能与 DNA 复制状态有关。
说明 M 期细胞中可能存在细胞有丝分裂促进因子:是 M 期细胞诱导 PCC 产生,暗示在
M 期细胞中可能存在一种诱导染色体凝缩因子。
1971,Yoshio Masui and Clement L Markert 用非洲爪蟾卵实验,发现成熟的卵细胞质中,
含有诱导卵母细胞成熟的物质,即 MPF。
Masui 和 Markert 分离出第Ⅳ期等待成熟的非洲爪蟾卵母细胞,并用孕酮进行体外刺激,
诱导卵母细胞成熟,然后进行细胞质移植实验,他们发现,在成熟的卵细胞的细胞质中
必然有一种物质可以诱导卵母细胞成熟,即成熟促进因子(MPF);
后来还证明,在成熟卵细胞中,MPF 已经存在,只需通过翻译后修饰即可转化为活性状
态的 MPF。
1988,JL Maller 实验室 MJ Lohka 以爪蟾为材料,分离得到 MPF,为 p32 和 p45 两种组
分蛋白, p32+p45→蛋白激酶活性→蛋白底物磷酸化。
故 MPF 是蛋白激酶
周期蛋白:
周期蛋白种类:多种类型,人 12 个以上,线虫 34 个
细胞周期内表达时期:G1 期,M 期等。
结构特点:周期蛋白均含有一段相当保守的氨基酸序列,为周期蛋白框 (cyclin box),可
介导 cyclin 与 CDK 结合,从而形成不同的 cyclin-CDK 复合体,表现出不同的 CDK 活性。
周期蛋白框(cyclin box):指所有周期蛋白中都存在的约由 100 个残基组成的相当保守
的氨基酸序列,其功能是介导周期蛋白与 CDK 结合。
PEST 序列:指 G1 期周期蛋白 cyclin D 的 C 端含有的一段特殊的 PEST 氨基酸序列( proline
(P), glutamic acid (E), serine (S), and threonine (T)),其功能可能与 G1 期周期蛋白的更新
(降解)有关。
Cyclin 的周期性变化:
CDK 是细胞周期运转的引擎分子,对细胞周期起着核心调控作用。不同种类的 cyclin 与
不同种类的 CDK 结合,构成不同活性的 CDK,在细胞周期具有不同的活性,对细胞周期
的不同时期进行调节。
癌基因:
1、癌基因 oncogene:是控制细胞生长和分裂的一类正常基因,其突变能引起正常细胞
发生癌变。
2、分类:两大类,源于病毒的 V-oncogene—指反转录病毒基因组里带有可使受感染宿
主细胞发生癌变的基因,如 src;源于细胞本身的,c-oncogene:指源于细胞的正常基因
突变产生,如 ras,故名原癌基因, 已识别的 100 多个
原癌基因存在于正常细胞中,控制细胞生长与增殖。往往是显性突变
癌基因起源于细胞,并普遍存在于许多生物基因组中。癌基因编码的蛋白质主要包括(1)
生长因子、(2)生长因子受体、(3)信号转导通路中的分子、(4)基因转录调控因
子、(5)细胞周期调控蛋白、(6)DNA 修复相关蛋白和(7)细胞凋亡蛋白等 7 大类
型。细胞信号转导是细胞增殖与分化过程的基本调控方式,而信号转导通路中蛋白因子
的突变是细胞癌变的主要原因。
抗癌基因(抑癌基因):是一类能够阻滞细胞周期进程的细胞周期负调控因子,通过阻
滞细胞周期进程,促进细胞凋亡等,抑制癌症的发生,如 p53,Rab 等
Rb 基因突变,即 Rb 失活,导致癌症易发,这类基因为抑癌基因。隐性突变。
p53 为抑癌基因,编码的基因可对细胞周期进程进行负调控。
细胞周期检查点:
DNA 损伤引起的细胞周期调控——S 期细胞周期检查点的作用。
DNA 复制不完成, S 期细胞周期检查点监控,CDK1 就无活性,因为 Cdc25 活性低,而
Weel 活性高,抑制 MPF 的活化
1、S 期内部检查点(intra-S phase checkpoint):指在 S 期内发生 DNA 损伤(DSB)时,
S 期内部检查点被激活,从而抑制复制起始点的启动使 DNA 复制速度减慢,S 期延长,
同时激活 DNA 修复和复制叉的恢复等机制。
2、 DNA 复制检查点(DNA replication checkpoint):指由于停滞的复制叉导致的 S 期的
延长。这 2 种检查点能够将细胞停滞在 S 期和 G2/M 期。当 DNA 复制叉阻断引发单链
DNA 时,ATR-CHK1 通路被激活;当 DNA 双链断裂时,ATM-CHK2 通路被激活。ATR/ATM
是 DNA 损伤信号感受因子,也是与 PI-3-K 同源的激酶,激活下游信号通路。CHK 即检查
点激酶。
S/G2/M 期转化与 DNA 复制检查点
第十四章 细胞分化与干细胞
细胞分化(cell differentation):在个体发育中,由一种相同细胞类型经细胞分裂后,
逐渐在形态,结构和功能上形成稳定性差异,产生不同的细胞类群的过程称为细胞分化. 分化细胞的特征:(1)持久性:细胞分化贯穿于生物体整个生命进程中,在胚胎时期达到
最大限度(2)稳定性:一般来说,分化了的细胞将一直保持分化后的状态,直至死亡(3)
普遍性:是生物界普遍存在的生命现象,是生物个体发育的基础.(4)不可逆性:细胞只
能从全能干细胞最终走向高度分化的体细胞,不能反向进行。(即全能性逐渐减小)
细胞分化的基因选择性表达:基因的选择性表达主要是由转录调节因子蛋白所启动。转
录因子蛋白的组合是影响细胞分化的主要的直接因素。一般来说,这种影响主要是胞外
信号及细胞微环境,同细胞内的信号转导调控网络相互作用,转录因子蛋白的表达受细
胞内外信号的精确调控。而胞外信号及细胞微环境又是通过信号转导调控网络来起作用。
在很多物种中,影响细胞分化的胞内因素,可以追溯到单细胞受精卵中细胞质的作用。
此外,外部的环境对某些物种的细胞分化乃至个体发育也会产生很大的影响
组合调控作用:组合调控:每种类型的细胞分化是由多种调控蛋白共同调控完成的,依
赖少量调控蛋白即可以启动众多细胞类型的基因表达。主导基因(mastergene):在细胞
分化启动过程中起决定作用的调控蛋白基因。生物学作用:借助于组合调控,一旦某种
主导基因产物与其它调控蛋白形成适当的调控蛋白组合,就可以将一种类型的细胞转化
成另一种类型的细胞,而且遵循类似的机制,甚至可以诱发整个器官的形成。
管家基因(house-keeping genes):管家基因是指所有细胞中均表达的一类基因,其产物
是对维持细胞基本生命活动所必需的;多在 S 早期复制,占 3%,转录起始区无 TATA box,
但有 GC 富集区,GC 岛。
组织特异性基因(tissue-specific genes),或称奢侈基因(luxury genes):是指不
同的细胞类型特异性表达的基因,其产物赋予各种类型细胞特异的形态结构特征与特异
的功能。调节基因产物用于调节组织特异性基因的表达,起激活或者起阻遏作用。
转分化(transdifferentiation):一种类型分化的细胞转变成另一种类型的分化细胞现
象称转分化转分化经历去分化(dedifferentiation)和再分化的过程. 去分化(dedifferentiation):又称脱分化,是指分化细胞失去其特有的结构与功能,
变成具有未分化细胞特征的过程。
细胞全能性(totipotency):是指细胞经分裂和分化后仍具有产生完整有机体的潜能或
特性。
细胞系(cell line):指原代细胞培养物经首次传代成功后所繁殖的细胞群体。也指
可长期连续传代的培养细胞。
细胞株(Cell Strain):通过选择法或克隆形成法从原代培养物或细胞系中获得具有
特殊性质或标志物的培养物称为细胞株。
原代细胞(primary culture cell):是指从机体取出后立即培养的细胞
传代细胞(sub culture cell):细胞群已经过数代培养,转到另一培养条件下培养的细
胞群。
多潜能干细胞:分化产生三个胚层中各种类型的细胞,并形成器官的一类干细胞,如胚
胎干细胞和生殖嵴干细胞。
多潜能性(pluripotency):高等动物细胞随着胚胎的发育,细胞逐渐丧失了发育成个体
的能力,仅具有分化成有限细胞类型及构建组织的潜能的现象。
诱导多能干细胞(induced pluripotent stem cell,iPS):通过导入外源基因的方法使
体细胞去分化形成多能干细胞,这类细胞谓之 iPS。由山中伸弥 Yamanaka 发现
干细胞(stem cell):是机体中能进行自我更新并具有多向分化潜能的一类细胞。干细胞
在个体发育、成体维持等生命过程中,具有关键和决定性作用。
多能干细胞(multipotent stem cells):具有分化成多种细胞类型或多种分化潜能的干
细胞。
第十五章 细胞衰老与细胞程序性死亡
细胞衰老(cell senescence):是指体外培养的正常细胞经过有限次数分裂后,停止分
裂,细胞形态和生理代谢活动发生显著改变的现象. 细胞衰老的主要特征:1、衰老细胞形态学特征:(1)外形变化:大而扁平(2)细胞核的
变化:体外培养的二倍体细胞,细胞核随着细胞分裂次数的增加增大;细胞核的核膜内
折(invagination)、染色质固缩化。(3)内质网:衰老动物内质网成分弥散性地分散
于核周胞质中,粗面内质网的总量呈减少趋势(4)线粒体:线粒体的数量随龄减少,而
其体积则随龄增大。(5)致密体的生成:即衰老细胞内由单层膜构成的,具有阳性磷
酸酶反应的结构。(6)膜系统的变化:衰老的细胞,其膜流动性降低、韧性减小;衰
老细胞间间隙连接及膜内颗粒的分布也发生变化。
衰老细胞的生物学特征(标志物):(1)生长停滞,细胞分裂不可逆的停止,不能通过添
加生长因子逆转;(2)若干细胞周期负调控因子如 p21,p53 表达上调(3)生化变化:
β-半乳糖甘酶活化,标志物(4)产生 lipofuscin 色素,标志物(5)衰老细胞端
粒长度明显减少(短于原有长度 50%)(6)出现衰老相关异染色质集中,主要使一些细胞
周期正向调控基因关闭(7)衰老特征分泌物,如炎性因子、金属蛋白酶等活性增加,
募集免疫细胞吞噬衰老细胞
3、分子水平的变化(1)DNA:复制与转录受到抑制,但也有个别基因会异常激活,端粒
DNA 丢失,线粒体 DNA 特异性缺失,DNA 氧化、断裂、缺失和交联,甲基化程度降低。
(2)RNA.mRNA 和 tRNA 含量降低。(3)蛋白质:合成下降,细胞内蛋白质发生糖基化、
氨甲酰化、脱氨基等修饰反应,导致蛋白质稳定性、抗原性,可消化性下降,自由基使
蛋白质肽断裂,交联而变性。氨基酸由左旋变为右旋。(4)酶分子:活性中心被氧化,
金属离子 Ca2+ Zn2+ Mg 2+、Fe2+等丢失,酶分子的二级结构,溶解度,等电点发生
改变,总的效应是酶失活。(5)脂类:不饱和脂肪酸被氧化,引起膜脂之间或与脂蛋白
之间交联,膜的流动性降低。
3、Hayflick 界限:细胞,至少是培养的细胞,不是不死的,而是有一定的寿命,它们
的增殖能力不是无限的,而是有一定的界限的现象。
细胞衰老的变化:(1)细胞水分减少,致使细胞脱水收缩,体积变小。(2)老年色素
积累,溶酶体功能低下,脂褐质积累,细胞效率降低出现衰老。(3)细胞核固缩,常染
色质减少。(4)细胞膜变厚,流动性随年龄增长而下降,物质转运及信息传递障碍,对
环境的应激能力迟钝。细胞在衰老过程中,其结构发生一系列的深刻变化,包括:①细
胞核增大,核膜内折,染色质固缩;②粗面内质网排列无序、趋于解体、总量减少;③
线粒体数量减少、体积增大、多囊体出现;④致密体产生并积累;⑤膜常处于呈凝胶相
或固相;⑥细胞间隙连接减少,组成间隙连接的膜内颗粒聚集体变小。
细胞衰老的机制:(一)复制性衰老(replicative senescence,RS):Harley 等 1991
发现体细胞染色体的端粒 DNA 会随细胞分裂次数增加而不断缩短。细胞 DNA 每复制一次
端粒就缩短一段,当缩短到一定程度至 Hayflick 点时,会启动 DNA 损伤检测点(DNA
damagecheckpoint),激活 p53,引起 p21 表达,导致不可逆地退出细胞周期,走向衰亡。
(二)胁迫诱导的早熟性衰老(SIPS)1、除了细胞内端粒缩短可以诱发复制衰老以外,
许多刺激因素,如离子辐射、过量的氧、乙醇和丝裂霉素 C 等非端粒 DNA 损伤,均能够
缩短细胞的复制寿命,促进细胞衰老。SIPS 与 RS 的机制相似。2、氧化性损伤学说:代
谢过程中产生的活性氧(ROS)成分(O2-,OH·,HzO2),对核酸、蛋白质和脂质造成损伤
并使线粒体 DNA 发生特异性突变,引发的氧化性损伤的积累,最终导致衰老。
细胞程序性死亡(Yprogrammed cell death,PCD):是指细胞的死亡是按照严格的遗传
机制的程序控制,是细胞主动性死亡,存在于所有细胞类型中。是维持生物体正常生长
发育及生命活动的必要条件。
细胞凋亡(apoptosis):是一种有序的或程序性的细胞死亡方式,是细胞接受某些特
定信号刺激后进行的正常生理应答反应,细胞凋亡过程具有典型的形态学和生化特征,
凋亡细胞最后以凋亡小体的形式被吞噬消化。
细胞凋亡的形态学变化:首先出现的是细胞体积缩小,连接消失,与周围的细胞脱离,
然后是细胞质密度增加,线粒体膜电位消失,通透性改变,释放细胞色素 C 到胞浆,核
质浓缩,核膜核仁破碎,DNA 降解成为约 180bp-200bp 片段;胞膜有小泡状形成,膜内
侧磷脂酰丝氨酸外翻到膜表面,胞膜结构仍然完整,最终可将凋亡细胞遗骸分割包裹为
几个凋亡小体,无内容物外溢,因此不引起周围的炎症反应,凋亡小体可迅速被周围专
职或非专职吞噬细胞吞噬。细胞凋亡呈现出特征性形态学变化,主要包括细胞皱缩、染
色质凝集、凋亡小体形成、细胞骨架解体等,其中以胞核的变化最为显著;细胞凋亡时
细胞的生化改变具有复杂性和多样性,包括 DNA 片段化、多种蛋白酶控制、胞浆 Ca2+
持续升高、pH 的变化、线粒体在细胞凋亡中起重要作用。
细胞凋亡的形态学特征:(1)染色质聚集、分块、位于核膜上,胞质凝缩,最后核断裂,
细胞通过出芽的方式形成许多凋亡小体;(2)凋亡小体内有结构完整的细胞器,还有凝
缩的染色体,可被邻近细胞吞噬消化,因始终有膜封闭,没有内溶物释放,故不会引起
炎症;
凋亡小体:凋亡过程中断裂的 DNA 或染色质与细胞其它内含物一起被反折的质膜包裹,
形成的圆形小体,凋亡小体可被邻近细胞吞噬。
细胞凋亡的生理意义:
(1)细胞凋亡对于多细胞生物个体发育的正常进行,稳态的保持具重要意义。(2)免
疫耐受性的形成,通过凋亡清除识别自身抗原的 T 细胞克隆,避免发生自身免疫疾病
(3)生理保护,肿瘤监控,以及抵御外界各种因素的干扰方面都起着非常关键的作用。
(4)凋亡是一种生理性的保护机制,可以清除机体多余、受损或危险的细胞,维持机
体的稳态,清除病原体等。
(5)癌症治疗,是肿瘤治疗的重要策略,使肿瘤细胞凋亡。
细胞凋亡的生物化学特征:
(1)凋亡细胞中仍需要合成一些蛋白质,蛋白质 ATP 合成继续,但是在坏死细胞中
ATP 和蛋白质合成受阻或终止;
(2)核酸内切酶活化,导致染色质 DNA 在核小体连接部位断裂,形成约 200bp 整数倍
的核酸片段,凝胶电泳图谱呈梯状,形成 DNA ladder;
(3)凋亡细胞 tTG(组织转谷氨酰胺酶 tissueTransglutaminase)积累;凋亡细胞质膜
PS 翻转;
12、细胞凋亡检测方法:
(1)形态学观测:染色法 DAPI,AO 等透射和扫描电镜观察,染色体固缩,凋亡小体
形成等。核酸染料染色程度高于染色质。
(2)DNA 电泳:DNA 片段就呈现出梯状条带,提取 DNA,EB 染色,观察 DNA 梯状条带,
大小呈现出 200bp 或其倍数 DNA ladders)
(3)TUNEL 测定法,即 DNA 断裂的原位末端标记法
(4)彗星电泳法(comet assay):细胞经裂解处理后,经单细胞电泳和荧光染色,凋
亡细胞由于 DNA 降解为片段状,泳动快,细胞核呈彗星状图案,未凋亡细胞核保持圆球
形。(5)流式细胞分析:根据凋亡细胞 DNA 断裂和丢失,采用碘化丙淀染色使 DNA 产生
红色荧光,用流式细胞仪检出凋亡的亚二倍体细胞,同时又能观察细胞的周期状态。(6)
检测细胞质膜从内侧翻转至外侧,使用特异性荧光探针检测
几个主要基因的作用:
P53:p53 是一种抑癌基因,其生物学功能是在 G1 期监视 DNA 的完整性。如有损伤,则
抑制细胞增殖,直到 DNA 修复完成。如果 DNA 不能被修复,则诱导其调亡,研究发
现丧失 p53 功能的小鼠对辐射诱导的调亡不敏感。缺失抑制凋亡,癌症高发!!!
Apaf-1:Apaf-1 被称为凋亡酶激活因子
-1(apoptoticprotease activating factor-1),在线虫中的同源物为 ced-4,在线粒
体参与的凋亡途径中具有重要作用。Apaf-1 含有 3 个不同的结构域:①CARD(caspase
recruitment domain)结构域,能募集 caspase-9;②ced-4 同源结构域,能结合
ATP/dATP;③C 端结构域,含有色氨酸/天冬氨酸重复序列,当细胞色素 c 结合到这一区
域后,能引起 Apaf-1 多聚化而激活。Apaf-1/细胞色素 c 复合体与 ATP/dATP 结合后,
Apaf-1 就可以通过其 CARD 结构域召集 caspase-9,形成凋亡体(apoptosome),激活
caspase-3,Apaf-1 启动 caspase 级联反应。缺失抑制凋亡!!!未缺失时促进细胞凋
亡。
Bcl-2:Bcl-2 为凋亡抑制基因是膜的整合蛋白,其功能相当于线虫中的 ced-9。根据功
能和结构可将 Bcl-2 基因家族分为两类(1)一类是抗凋亡的(anti-apoptotic),如:
Bcl-2、Bcl-xl、Bcl-w、Mcl-1;(2)一类是促进凋亡的(pro-apoptotic),如:Bax、
Bak Bad、Bid、Bim,在促凋亡蛋白中还有一类仅含 BH3 结构,如 Bid、Bad。
(SoBH3-only domain!),BH3 主要定位于线粒体外膜,它拮抗促凋亡蛋白的功能。缺失
促进凋亡!!!,未缺失时抑制细胞凋亡 caspase-9:裂解核纤层蛋白 B,导致细胞核
形成凋亡小体·9 裂解 DNase 结合蛋白,使 DNase 释放,降解 DNA 形成 DNA Ladder;
裂解参与细胞连接或附着的骨架和其他蛋白,使凋亡细胞皱缩、脱落,便于细胞吞噬,
导致膜脂 PS 重排,便于吞噬细胞识别并吞噬; caspase 还可切割黏着斑激酶,破坏细
胞黏着,使凋亡细胞与邻近细胞脱离;失活 DNA 修复相关酶类,促进 DNA 降解等。Caspase
突变或缺失,失去凋亡能力!!!
ATM:ATM(ataxia telangiectasia-mutated gene 是与 DNA 损伤检验有关的一个重要
基因。最早发现于毛细血管扩张性共济失调症患者,人类中大约有 1%的人是 ATM 缺失的
杂合子,表现出对电离辐射敏感和易患癌症。正常细胞经放射处理后,DNA 损伤会激
活修复机制,如 DNA 不能修复则诱导细胞凋亡。ATM 是 DNA 损伤检验点的一个重要的蛋
白激酶
细胞坏死(necrosis):被动的细胞死亡,是细胞受到意外损伤,如极端的物理、化学
或严重病理刺激下导致的细胞死亡。
细胞自噬:(1)概念:正常细胞为维持细胞内环境的动态平衡,对功能失常或不需要
的细胞结构通过形成双层膜包裹的自噬小体,由溶酶体进行降解的过程,标志是细胞内
双层膜包裹的自噬小体(2)降解途径:蛋白质:泛素-蛋白酶途径;细胞器及长寿命蛋
白-细胞自噬途径,由溶酶体进行降解(3)降解状态:生理状态:营养危机或胚胎发育,
降解自身细胞产生能量,导致细胞死亡,同时清除一些不需要的细胞。(4)调控:PI3K
途径可抑制细胞自噬,而营养不良将导致 PI3K 下游 AKT 失活,抑制 Tor 蛋白,自噬相
关蛋白 Atg 活化,细胞死亡
细胞凋亡的分子机制:
1、诱导凋亡的因子:(1)物理性因子,包括射线(紫外线,y 射线等),较温和的温度
刺激(如热激,冷激)等;(2)化学因子,有毒物质,活性氧(ROS)基团;(3)生物
因子,DNA 和蛋白质合成的抑制剂,激素,细胞生长因子,肿瘤坏死因子α(TNFa,抗
Fas/Apo-1/CD95 抗体等;(4)病菌感染;(5)生理性凋亡,维持机体稳态。2、动物
细胞的凋亡过程:(1)接收凋亡信号; (2)凋亡相关分子的活化; (3)凋亡的
执行;(4)凋亡细胞的清除几个阶段。
3、细胞凋亡的途径主要包括 2 条:
(1) Caspase (cysteine aspartic acid specificprotease)依赖性细胞凋亡途
径;(2)非 Caspase 依赖性细胞凋亡途径。当细胞受到凋亡信号的刺激时,这 2 条途径
一般能够同时被激活。
4、caspases(1)caspase Cysteine aspartic acid specific protease)是存在
于细胞质中的半胱氨酸蛋白酶,其活性位点包含 Cys 残基,特异性水解靶蛋白 Asp 基端
肽键,故又称为天冬氨酸特异性的半胱氨酸蛋白水解酶,也叫半胱天冬酶。根据 caspase
在凋亡中的作用,可以把 caspase 分为 (a)凋亡起始者(apoptotic initiator)如
caspase-2,8 , 9 , 1011 和 (b) 凋 亡 的 执 行 者 (apoptotic executioner) ,
如 caspase-3,6,7O 起始者对执行者前体进行切割激活,后者再切割相关蛋白,促使凋
亡。
5、细胞凋亡过程包括激活期和执行期两个过程激活期细胞应答死亡信号,起始
caspase 活化,执行期执行 caspase 活化,执行细胞死亡程序。
(1)Caspase 的活化机制,caspase 在细胞质基质中以无活性酶原形式存在,凋亡信号
刺激可使 caspase 酶原(procaspase)之特异的 Asp 位点被切割,形成大小两个亚基
的异二聚体,表现出 caspase 的活性。由激活的起始 caspase 招募执行 caspase 酶原,
并对其进行激活,产生具有活性的执行caspase,执行caspase 可作用于多种底物(>280
种),促使细胞呈现出凋亡的形态和分子生物学特征。(2)起始者 caspases 的同性活化
(homo-activation):即同一种 caspases 酶原分子彼此结合或与接头蛋白结合形成
复合物,此间构象改变被活化,进而彼此切割产生有 2 个活性的二聚体形式。
在 起 始 者 caspase 中 , caspase-8 和 -10 含 有 串 联 重 复 的 死 亡 效 应 结 构 域
(death effectordomain,DED);而 caspase-2 和 -9 含有 caspase 募集结构域
(caspase recruitmentdomain,CARD)。这 2 种结构域存在于 caspase 酶原分子和一些
凋亡相关的接头蛋白分子中,通过结构域之间的聚合,caspases 能够彼此结合或与接头
蛋白结合,被招募到上游信号复合物中而发生同性活化。(3)效应者 caspases 的异性活
化,即已活化的起始者 caspases 招募效应者 caspases 酶原分子后,对其进行切割,
产生具有活性的效应者 caspases。活化的效应者 caspases 切割细胞中重要的结构蛋白
(如支架蛋白)和调节蛋白(如信号蛋白、TFs 和周期蛋白等),导致细胞凋亡。(4)效应者
caspases 底物:约 280 种,可分为活化和失活 2 大类。
被 效 应 者 caspase 活 化 的 代 表 底 物 — Caspase 激 活 的 DNA 酶
(Caspase activated Dnase,CAD。细胞凋亡程序启动后,活化的效应者 Caspase-3 降
解 ICAD 使 CAD 释放出来并在核小体间切割 DNA,形成间隔 200bp 或其整倍数的 DNA
片段,琼脂糖电泳时产生凋亡标志性特征的梯状条带。被效应者 caspase 失活的代表底
物:众多被效应者 caspase 失活的代表底物都在维护细胞正常状态中发挥关键作用,如
多聚(ADP-核糖)聚合酶(PARP)、黏着斑激酶(FAK)和核纤层蛋白等。
第十六章 细胞的社会联系
通信连接(communicating junction):介导相邻细胞间的物质转运、化学或电信号的
传递,主要包括动物细胞间的间隙连接、神经元之间或神经元与效应细胞之间的化学突
触和植物细胞间的胞间连丝。以连接子为单位形成的连接方式。连接子蛋白的一级结构
都比较保守,并有相的抗原性。连接子蛋白具有 4 个α-螺旋的跨膜区,是该蛋白家族
最保守的区域。不同类型细胞表达不同的连接子蛋白。功能:
(1)代谢耦联:允许小分子代谢物和信号分子通过 <1k),促使细胞间代谢耦联,协
调细胞群体功能; Caz+通透性
(2)在神经冲动信息传递过程中的作用:电突触(electronicjunction)快速实现细胞
间信号通讯;间隙连接调节和修饰相互独立的神经元群的行为,协调心肌细胞平滑肌细
胞收缩。
(3) 间隙连接在早期胚胎发育和细胞分化过程中的作用,胚胎发育中细胞间的偶联提供
信号物质的通路,影响其分化。肿瘤细胞之间间隙连接明显减少或消失,间隙联接类似
“肿瘤抑制因子”。
(4)辐射损伤信号传导:旁效应
胞间连丝(plasmodesma):高等植物细胞间,由相邻细胞的细胞质膜共同组成直径为
20-40nm 的管状结构,中央是由 SER 延伸形成链样管,在链样管和管状质膜之间是由细
胞液构成的环孔。除极少数特化的细胞,外国高等植物细胞之间,通过胞间连丝相互连
接,完成细胞间的信号通络。胞间连丝可在植物细胞胞质分裂中细胞板形成后即产生,
也可在非姐妹细胞之间形成,而且还可以通过修饰改变它的结构和运输功能。
胞间连丝的功能:实现细胞间由信号介导的物质有择性的转运;实现细胞间的电传导;
在发育过程中,胞间连丝结构的改变可以调节植物细胞间的物质运输。
