
回顾:2022年的恐龙新种(上)
大家好,我是黑莓,我们又见面了。
2022已经过半,就让我们简单地整理回顾一下上半年命名的那些非鸟恐龙吧。(去年讲过的例如玉溪龙和迈普龙这里不再重复讲)

匿迹多年的外来者
2月7号,期刊《Nature Ecology & Evolution》刊登了对西班牙新种泰坦巨龙类的研究,来自西班牙的科学家命名了库氏匿踪龙(Abditosaurus kuehnei),属名是指从它被发现到现在已经被搁置了近60年。匿踪龙的化石出土于特雷姆群(Tremp Group),正模标本包含大量颈椎椎体、背椎以及肋骨、部分四肢以及部分骨盆。
匿踪龙属于泰坦巨龙类,不同于当时欧洲群岛的诸多矮化泰坦巨龙类,它的体型要更大一些,并且肱骨要更加粗壮。系统发育分析显示:匿踪龙属于萨尔塔龙亚科,与它更加亲近的那些来自非洲和南美的谱系而非欧洲的谱系,这表明它可能是一个从冈瓦纳大陆迁徙到欧洲群岛的外来客。

骨组织研究显示:匿踪龙的附肢骨骼缺乏重组的薄层骨,这个结构很可能与减小新陈代谢速率和体型有关,被认为是检测蜥脚类恐龙岛屿矮化的工具之一。而缺乏这个结构,表明匿踪龙本身可能并没有出现与岛屿矮化现象相关的新陈代谢变化。这也是支持匿踪龙属于其它大陆外来者而非欧洲原生谱系的证据之一。

至于匿踪龙迁徙的途径,科学家认为:在马斯特里赫特期早期(70.6Ma前后),海侵停止,海平面有所下降,连接欧洲和冈瓦纳的陆桥和群岛重新出现,不少物种得以从非洲迁徙到伊比利亚-阿摩力克群岛。我们有理由相信,这种外来客会对当时的岛屿生态系统产生非常巨大的影响,蜥脚类恐龙的组成也会发生十分明显的变化。

袖珍将军
2月10号,《Journal of Vertebrate Paleontology》上刊登了一种新的小型阿贝力龙科成员,来自英国和阿根廷的学者将其命名为奥氏圭梅斯龙(Guemesia ochoai)。圭梅斯龙生存于坎潘期的阿根廷西北部,化石出土于洛斯布罗基托斯组(Los Blanquitos Formation),正模标本IBIGEO-P 103是一个几乎完整的脑壳。
圭梅斯龙的体型很小,可以说是在目前已命名的阿贝力龙科中最小的一批,但通过矮而窄的顶骨结节推测它可能是个未成年个体,这点还需要更多的材料来证明。圭梅斯龙拥有许多全新的形态特征,它的额骨中线上有几个大而浅的凹坑,额骨眶区有一个明显的空腔。系统发育分析显示:圭梅斯龙属于衍化的短吻龙类。


关于圭梅斯龙这两个自有衍征的作用,科学家认为,圭梅斯龙额骨上的凹坑可能有着感光感热的作用,但因为在脑部扫描中并没有发现类似的神经管道结构,所以它的作用机理尚不能确定;而额骨眶区的空腔既可能是一个血窦结构,用于供血和温度调节,也可能是一个气腔化的腔室,用于颅骨减重。

在晚白垩世时期,海侵规模增大,南美被分割成两块大陆,两侧陆地脊椎动物的联系减少。而位于阿根廷西北角的圭梅斯龙与其它阿贝力龙类展现出的差异或许正是长期地理隔离的结果。
伊比利亚的棘龙
早白垩世的欧洲作为棘龙科的演化焦点之一,存在着许多不为人知的新物种,在2月16号《PLOS ONE》上发布的论文中,来自葡萄牙的科学家基于新的材料将曾经被归于沃氏重爪龙(Baryonyx walkeri)的ML 1190建立新属种并命名为纳氏伊比利亚棘龙(Iberospinus natarioi)。伊比利亚棘龙生存于早白垩世的葡萄牙,化石出土于帕波塞可组(Papo Seco Formation),正模标本ML 1190包含部分下颌、椎骨、肩带和骨盆。

伊比利亚棘龙拥有一些独立于其它棘龙科的特征例如齿骨仅有一个麦氏孔、笔直的齿骨腹侧缘、中段尾椎的板层结构等等。系统发育分析显示:它属于位于棘龙科的基干位置,但一些特征还是将它指向重爪龙亚科。

虽然伊比利亚棘龙是比较基干的棘龙科成员,但它已经有了一些亲水的特征,齿骨拥有发达的神经管道,这可能表明它拥有不错的触觉系统;牙槽中存在两颗替换齿,表明它可以时刻保持牙齿的功能完善;椎体前表面的模式接近于棘龙但整体的椎体形态依旧与普通兽脚类类似,表明它处在一个进化的过程中;脚趾的结构介于其它坚尾龙类和棘龙之间也表明伊比利亚棘龙并没有像棘龙那样过着极端的水生生活。


“蛋蜥蜴”
2月16号,期刊《Cretaceous Research》上发表了一种全新的阿尔瓦雷斯龙类,来自俄罗斯的科学家命名了阿氏蛋龙(Ondogurvel alifanovi)。蛋龙生存于晚白垩世的蒙古,化石出土于巴伦戈约特组(Barungoyot Formation),正模标本PIN 5838/1包含部分椎骨、骨盆和肢骨。

蛋龙属于阿尔瓦雷斯龙科,它第二和第四跖骨的连接处完全愈合。系统发育分析显示:它属于小驰龙亚科,与第二第四跖骨接近愈合的西峡爪龙和游光爪龙接近。

在小驰龙亚科中,腕掌骨的形态主要有两种,而卵岩龙属于第一种,即三四掌骨的关节面位于腕掌骨侧面,较小但仍保留有功能的三四指的弯曲面几乎垂直于第二指的弯曲面。除了蛋龙,单爪龙也是这样的腕掌骨形态,而以临河爪龙为代表的物种则是第二类,即第四掌骨关节面缺失,第三掌骨的关节面位于腕掌骨的远端,靠近第二掌骨的远端髁。参考系统发育分析,或许表明在小驰龙亚科当中腕掌骨形态的差异也代表着不同谱系之间的差异。

赤色高原的萨满
在2月24号《Journal of Systematic Palaeontology》发表的论文上,来自阿根廷的科学家描述了晚白垩世两种新的鸭嘴龙类,其中一个就是萨满红壤龙(Kelumapusaura machi)。红壤龙生存于晚白垩世的阿根廷,化石出土于艾伦组(Allen Formation),正模标本MPCN-PV-807包含部分头骨和椎骨还有极少量的腹膜肋,而除了正模,它还有一些其它的标本。红壤龙的标本与去年的深潭龙出土于同一个骨床,包含多个个体,其中最大的MPCN-PV-808(部分右齿骨)可以达到近9米。

作为一个新的鸭嘴龙属种,红壤龙有许多其它鸭嘴龙类没有的特征,它的第一骶椎关节面明显向前上方倾斜,下颌联合处有一个明显的亚三角形突起,上颌骨低矮、存在一排小孔。系统发育分析显示:红壤龙属于栉龙亚科的小贵族龙族(Kritosaurini)。


不为人知的全新支系
在描述红壤龙的同时,科学家还建立了一个新属种——南方鹜龙(Huallasaurus australis)。鹜龙也生存于晚白垩世的阿根廷,但它的化石出土于洛斯阿拉米托斯组(Los Alamitos),正模标本MACN-PV 2包含部分头骨、椎骨、肩带、腰带和附肢。

鹜龙曾经被归于小贵族龙属,而后又被归于科氏独孤龙(Scernosaurus koerneri),但在之后的研究中,科学家发现独孤龙的髋臼后突的形态更接近于埃德蒙顿龙、昆杜尔龙等北方类群,但马普切鸭龙的髋臼不仅更接近其它的巴塔哥尼亚鸭嘴龙,更是有着独特的特征。在今年的新研究中,科学家指出了它的自有特征组合,将其单独列为新属。

系统发育分析显示:鹜龙和红壤龙均属于栉龙亚科的小贵族龙族,但它们同其它的南美栉龙类一起构成了单独的南美分支,平行于小贵族龙、格里芬龙所属的北美分支。这样的结果表明南美的鸭嘴龙科,至少是栉龙亚科并非是我们想象中罕见的流浪者,而是已经在南美开枝散叶,进行了辐射演化,并且可能是晚白垩世巴塔哥尼亚动物群的关键组成部分,只不过由于保存和研究问题一直没能够得到重视。
帝王和女王?
3月1号,由格瑞戈里•S•保罗撰写的霸王龙属分种论文被刊登在了期刊《Evolutionary Biology》上。保罗跟据齿骨前端牙齿的粗壮度和股骨粗壮度将霸王龙属(Tyrannosaurus)分为三个种:模式种同时也是态模2的君王霸王龙(Tyrannosaurus rex)普遍粗壮,股骨长与股骨周长的比值≤2.4,齿骨前的薄切齿只有一颗;态模1帝王霸王龙(Tyrannosaurus imperator)同样粗壮,股骨长与股骨周长的比值也≤2.4,但齿骨前有两颗薄切齿;态模3女王霸王龙(Tyrannosaurus regina)比较纤细,股骨长与股骨周长的比值>2.4,而齿骨前的薄切齿只有一颗。
保罗认为:这三者之间可能存在某种演化关系:年代最早的“粗壮型”帝王霸王龙演化出了年代较晚的君王霸王龙和女王霸王龙,在这期间,霸王龙的齿骨前的薄切齿数量减少,并且分化出了“粗壮型”君王霸王龙和“纤细型”女王霸王龙。
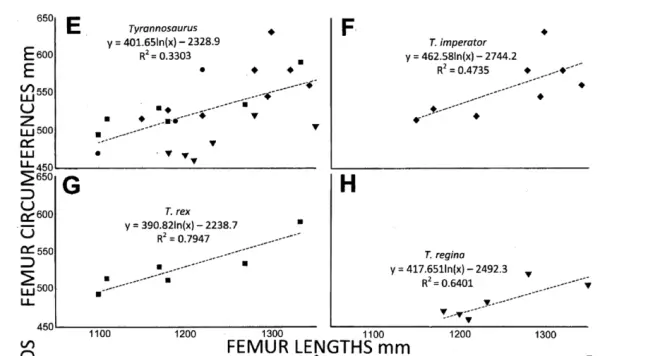
但话说回来,这种分法就一定是正确的吗?显然不是,保罗在论文中三分霸王龙的依据太少,仅跟据股骨粗壮度和齿骨前端牙齿将霸王龙分为三个不同的种显然太过武断,更不用说这种分化导致有不少的霸王龙标本无法被确切的归入这三个种中的任何一个。如果真的想要将不同类型的霸王龙区分开来,个人认为只需要在详细的形态学区分的基础上分出几个大类型即可,而过于简单粗暴的建立不同的态模标本不仅有失严谨,后续的研究也会因为必须考虑到它们的区别而花费大量额外的笔墨去把简单的东西复杂化,这是得不偿失的。

原始的巴山剑龙
时隔多年,来自国内的几位学者描述了国内剑龙类的一个新属种,并在3月3日由期刊《Journal of Vertebrate Paleontology》发布,他们将其命名为元始巴山龙(Bashanosaurus primitivus)。巴山龙生存于中侏罗世的中国,化石出土于沙溪庙组(Shaximiao Formation)下段,正模标本CLGPR V00006-1包含一些椎骨、肩带、附肢和骨甲。

巴山龙是一种生存于巴柔期的剑龙类,肩峰突的发育情况以及股骨第四转子的位置证明它是一个全新的物种。科学家通过其股骨推算得其体长约为2.8米,基于其肩胛乌喙骨以及胫腓骨未完全愈合的情况,它可能并没有达到成年;但其股骨转子以及肩胛骨、乌喙骨本身的情况更接近于成年的剑龙。总之,作为普遍不好探究生长发育阶段状态的剑龙类成员,目前判断巴山龙的生长发育情况是比较困难的。

系统发育分析显示:巴山龙是一种十分基干的剑龙下目成员,并且其它来自沙溪庙组的剑龙类例如华阳龙、重庆龙、巨刺龙等也都处于较基干的位置,这表明亚洲是剑龙类起源和发展的一个重要地点。

古老的中亚之爪
在蛋龙的论文中,科学家提及到土仑期乌兹别克斯坦贝斯克提组(Bissekty Formation)也有着阿尔瓦雷斯龙类,而在3月10号《Journal of Vertebrate Paleontology》发表的论文中,蛋龙的第一作者和另一位美国学者一起将贝斯克提组的阿尔瓦雷斯龙类命名为古老扎拉爪龙(Dzharaonyx eski)。扎拉爪龙的正模ZIN PH 2619/16是一个肱骨,但它还包括了很多其它部位的骨骼,包括椎骨、尺骨、腕掌骨、跖骨等等。

扎拉爪龙是一种阿尔瓦雷斯龙科成员,其后凹的后段背椎和神经弓上有纵向嵴线的前凹尾椎都证明它是一个全新的物种。系统发育分析显示:它属于小驰龙亚科,并且形态学特征也支持这一研究。

扎拉爪龙生存于土仑期,年代要明显早于其它大部分生存于亚洲的小驰龙类,这表明小驰龙亚科至少在土仑期就开始演化了,而亚洲很可能就是小驰龙亚科的发源地。

海岸旁的大爪子
5月3号,《Scientific Reports》刊登了一种来自日本的新种镰刀龙类,几位日本的科学家命名了日本滨镰龙(Paralitherizinosaurus japonicus)。滨镰龙生存于坎潘期,化石出土于无畏组(Osoushinai Formation),正模标本NMV-52包括一个残破脊椎和部分右手。滨镰龙是一种镰刀龙科的成员,最初描述时科学家只将它归类为手盗龙类,但最新的研究证明它是一种镰刀龙类。系统发育分析显示:它与镰刀龙、肃州龙以及贝斯克缇组的未定种镰刀龙类关系较近。

从结构上来看,滨镰龙的指爪与镰刀龙的十分类似,都有变细的倾向以及变小的屈肌结节,这或许会使得它们屈指的力量变弱;同时,它们发达的腹突也会限制屈指的幅度,因此像滨镰龙和镰刀龙这样的衍化镰刀龙类的手很可能并不会像那些基础的镰刀龙类那样拥有更多的功能而是主要侧重于勾住树枝,将植物送到离嘴边更近的位置。


作为日本最晚的镰刀龙类成员,滨镰龙使得镰刀龙类谱系在日本有着更长久的分布,同时大量的海生动物化石证明无畏组是一个沿海的环境,也就是说滨镰龙是一种典型的沿海生存的镰刀龙类。
上半年的非鸟恐龙新种总结到这里就结束了,总体来说,相比去年,今年的发力显然有些不足,但幸运的是:在我写完这篇文章不到一个小时后,藏在迷雾中的“坎鲨”就作为巨型米拉西斯龙现世,而下半年还会带来什么惊喜,那就让我们拭目以待吧。
