
你的NMN可能白吃了!NAD+消耗飙升成衰老罪魁祸首

NAD+作为机体必需辅酶,又因其含量随衰老下降,一直都是抗衰领域的香饽饽,其前体NMN类物质甚至还有了“长寿药”的美誉。
然而,NAD+为何伴随衰老下降却未有定论:究竟是合成不够,还是用得太多?
对此,一项发表于顶尖杂志Cell子刊的研究发现:导致老年个体NAD+不足的根本原因与NAD+合成无关,真相是NAD+的消耗效率大幅跃升!而抗衰“万金油”热量限制能显著降低NAD+消耗[1]。
本周日(12月10日),本文通讯作者宾夕法尼亚大学教授Joseph Baur将来到TIMEPIE第四届衰老干预论坛现场,就“线粒体NAD+代谢”做重磅分享。
推荐阅读 :


温故知新,NAD+代谢回顾
说起NAD+代谢途径及其前体,好似真能算上老生常谈,领域内的初步研究也已较为清晰,简单一图带你快速复习下👇
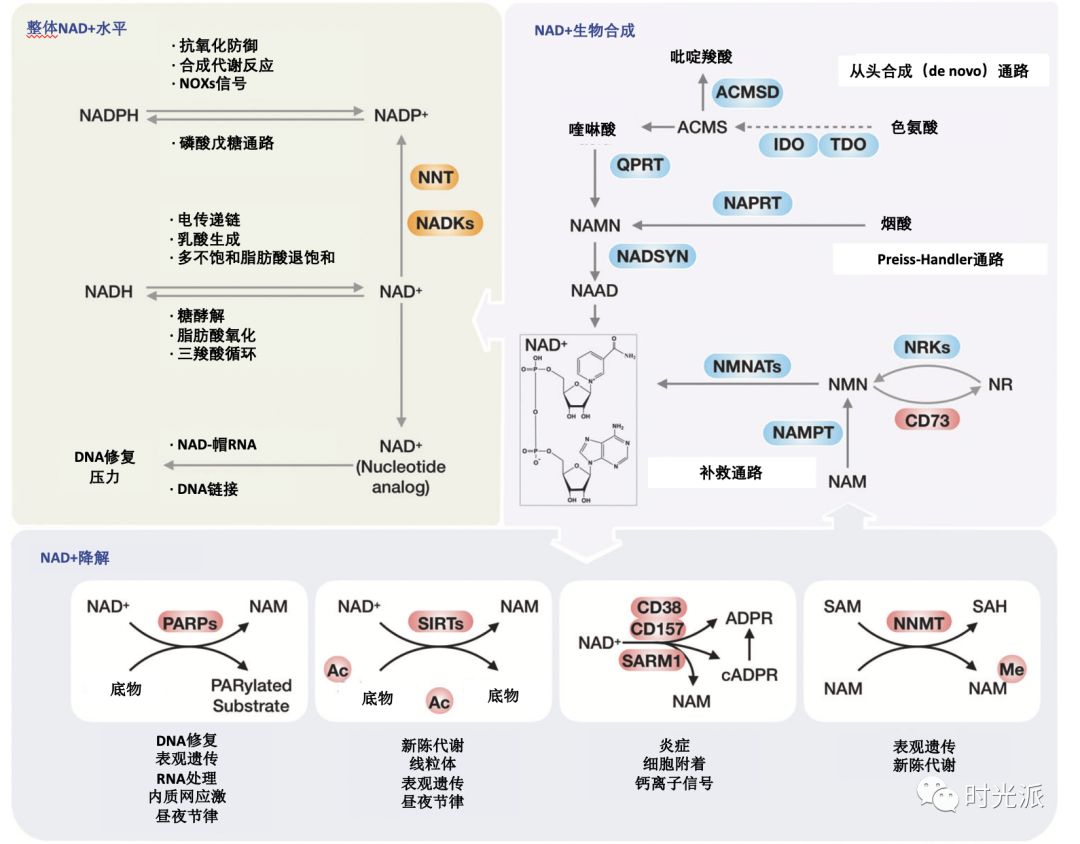
图注:NAD+主要代谢与生理功能总览源[2],汉化
NAD+消耗:
被多种关键酶作为底物利用,主要包括SIRTs、PARPs、CD38/CD157以及SARM1。其中,CD38和PARP1是细胞内最主要的NAD+使用者。
NAD+合成:
主要合成途径有三条,分别是经由色氨酸的从头合成通路、以烟酸(NA)为前体的Preiss-Handler通路以及通过烟酰胺(NAM)、NMN的补救通路。生物体多数组织中NAD+合成较依赖补救通路[3]。
不断被消耗,又不断重新合成,但这样“有来有往”的平衡终还是要被岁月打破,生物体NAD+含量随年龄增长而下降,已是学界公认事实[4, 5]。本次研究也同样发现,衰老促使小鼠体内多组织NAD+出现下降。
同时,研究人员还发现,除NMN外,多数NAD+循环前体的含量并未明显下降。言下之意,衰老并没拿着物质合成的生产原料“开刀”,NAD+真没到“穷得揭不开锅”的境地。
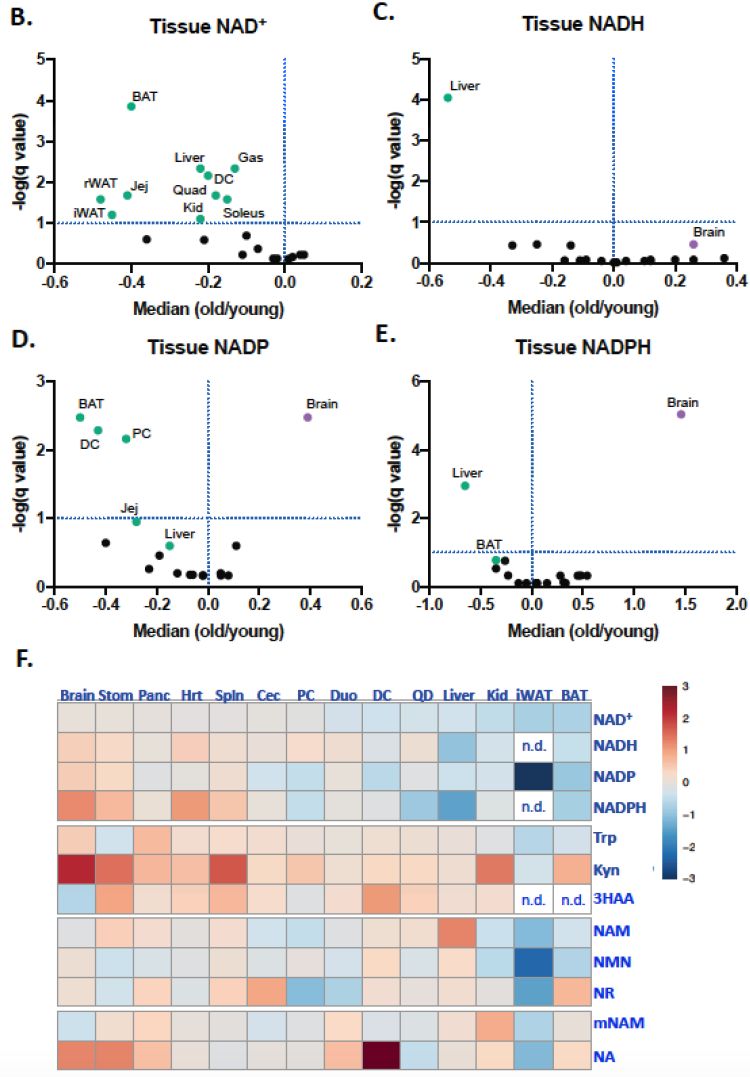
图注:小鼠体内NAD+及其多个前体含量随衰老的变化

颠覆认知!
NAD+下行并非合成不足,竟是消耗飙升
生产原料充足,NAD+却还是不够用,究竟是合成过程出了岔子,还是后端不加节制,消耗过快?要解答这个问题,还得一条条通路逐个量化、综合分析。
#No.1
从头合成通路
为了清晰追踪、量化NAD+关联过程,此次研究选择了堪称“最强追踪器”的同位素标记技术,给前端原料(色氨酸)贴上“特殊标签”,实时监测色氨酸、NAM及中间代谢产物的动态变化。
然而,有些令人意外的是,年轻和老年小鼠组织内被标记的NAD+、NAM表现出相似变化规律,无论是绝对含量,还是组织、时间差异,都与年龄因素无关。
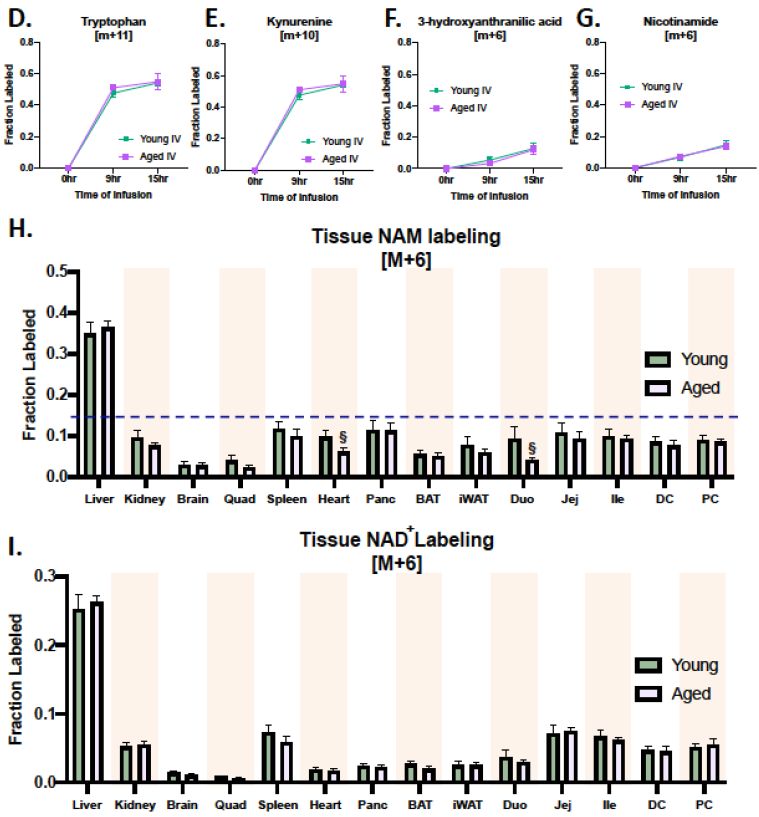
图注:从头合成通路中,NAD+的合成及该通路释放NAM含量未随年龄增长而改变
#No.2
补救通路
如法炮制,研究人员再次向小鼠体内注入标记NAM,观测经由补救通路的NAD+动态规律。24小时后,年轻与老年鼠体内NAD+水平均得到提升,且通量无明显差异,相比之下,衰老小鼠体内被探测到更高比例的标记NAD+。
这表明,老年鼠体内NAD+合成的绝对通量并未因年龄增长而改变,但消耗速率却在显著上升,这也与NAD+多种消耗酶活性随衰老增加的规律一致[3, 5]。
这道理就仿佛是:随着工龄增长,虽然收入并不见少,但奈何花销实在过大,到了着急用钱时,也只能看着存款余额黯然神伤。
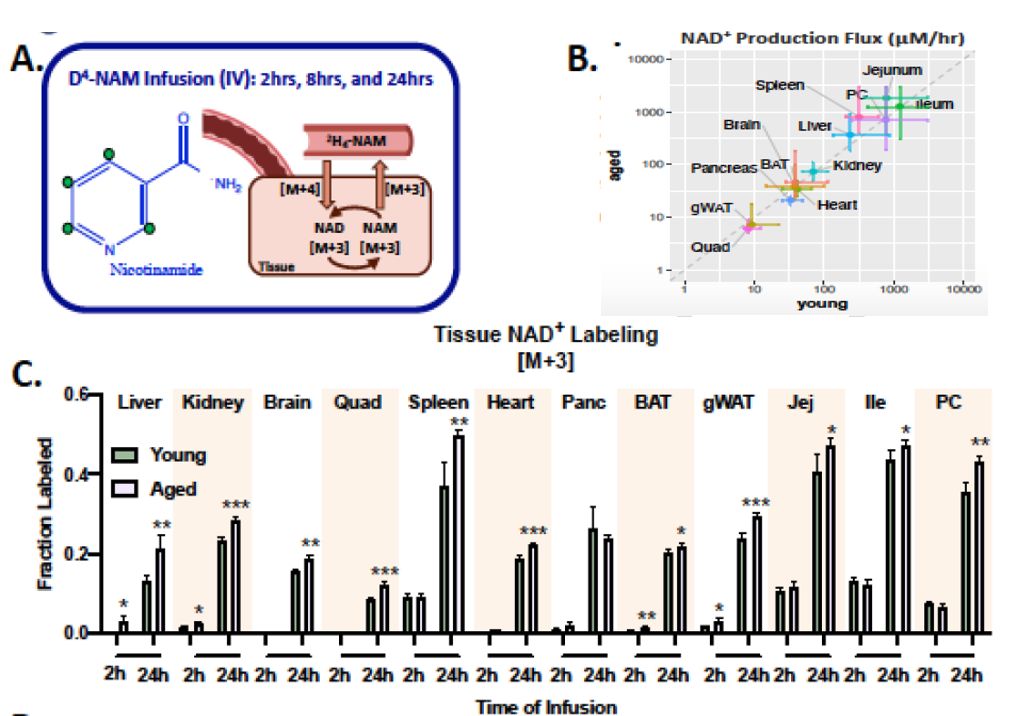
图注:老年小鼠体内NAD+合成未受到显著影响,但消耗明显增加

热量限制虽迟未缺,
大幅降低NAD+周转!
万万没想到,相比于“开源”,原来“节流”才是NAD+的稳态真谛,不过,想降低NAD+消耗,我们还能依靠谁?
这不,号称“已知的最强抗衰干预措施”[6]热量限制(CR)虽迟未缺。CR通过上调NAD+代谢的限速酶NAMPT[7],并调控NAD+重要消耗酶(SIRTs、CD38与PARPs)活性,进而降低DNA损伤,让NAD+水平远离衰老威胁[8, 9],可谓天长地久,多重保护。
而本次研究借助同位素“度量之手”,发现终生CR能让小鼠在变成“老鼠”时,体内各组织的NAD+稳态被极大改善,如肝脏NAD+水平得到恢复;白色脂肪组织(生物体中的“好脂肪”)的NAD+浓度“越活越年轻”,打破了衰老降低魔咒。
而这一成就还得归功于CR“稳住”NAD+“基本盘”,降低了衰老小鼠体内NAD+消耗——相比较对照组,CR组老年鼠体内被标记NAD+的比例大大降低,这意味原有NAD+的消耗被减缓,重新合成的NAD+较少。
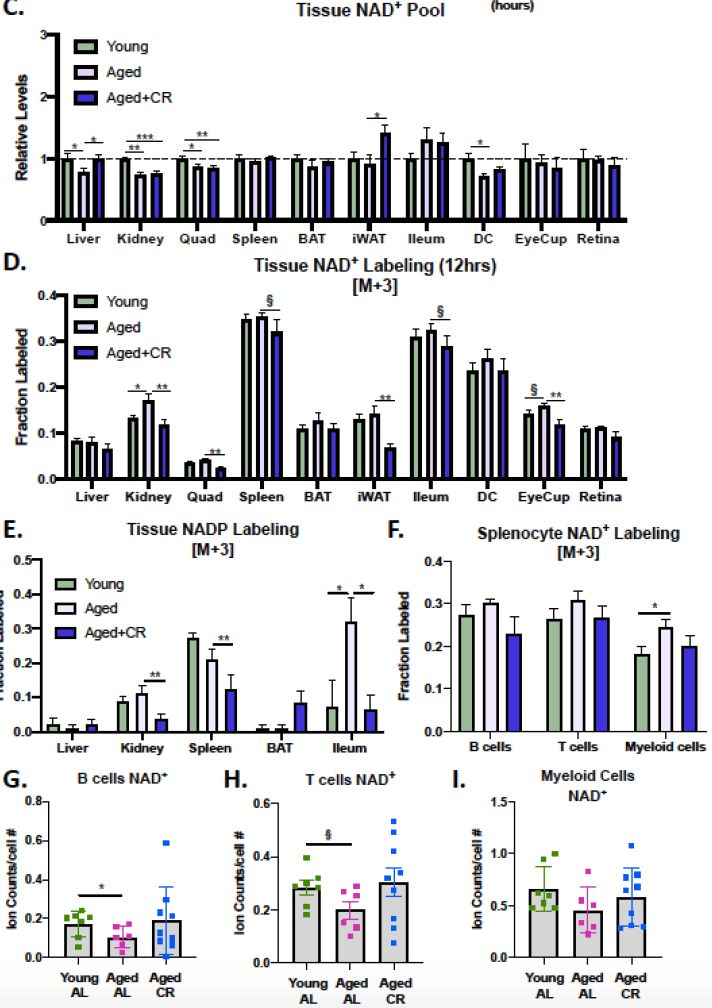
图注:热量限制适度增加了NAD+水平,并减少多数组织中NAD+的消耗
此外,CR似乎也让“动力工厂”线粒体变得更加年轻态。除了降低线粒体内的NAD+消耗,更是在肝脏“NAD+危机”到来之前(组织NAD+下降最终会破坏线粒体内NAD+稳态,但有一定延迟效应[10]),率先排兵布阵,守卫健康堡垒,适度提升了肝脏内NAD+的水平。
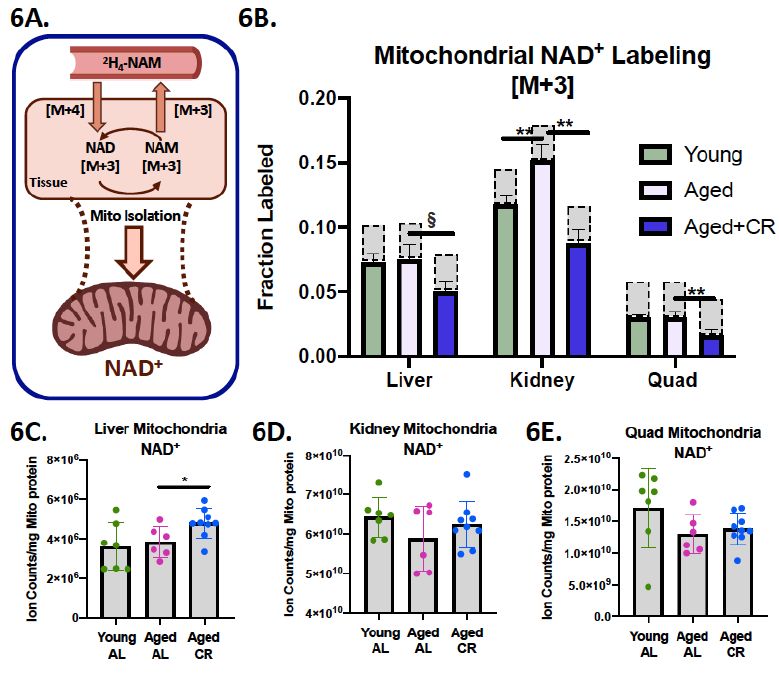
图注:线粒体NAD+代谢较机体组织更慢,CR能更好维持NAD+稳态
(不得不再次感叹,CR真的宇宙级yyds!!)
TIMEPIE点评
不同于主流观点,认为NAD+水平下降的主要原因是衰老影响合成,本次研究在量化生物体多组织NAD+代谢通量后,提出年龄增长导致的消耗飙升才是“拉扯”NAD+水平的罪魁祸首。而此次研究利用同位素技术,全面展现了多组织NAD+代谢“足迹”,并直接测量与年龄相关的变化,更是将NAD+的机制研究往前推进一大步。
“闪闪发光”的同位素信号告诉我们,衰老的确让机体利用NAD+前体的能力下降,因此,及时补充利用效率较好的前体物质,对稳定NAD+合成水平大有意义。但想要从根本上去遏制NAD+因年龄增长的变化,可能还是得再想想办法“堵住口袋底部的破洞”。
本周日(12月10日),本文通讯作者宾夕法尼亚大学教授Joseph Baur将来到TIMEPIE第四届衰老干预论坛现场,就“线粒体NAD+代谢”做重磅分享。
—— TIMEPIE ——
号外🔈千人”长寿盛会“【倒计时5天】!30+全球抗衰专家将于12月9日-10日空降上海,演讲主题涵盖最新衰老干预进展、行业动态、前沿技术等,来线下面对面对话学者大咖。(感兴趣的读者可点下方海报购买,如有疑问,加Vx:fudan246。)
参考文献
[1] McReynolds, M. R., Chellappa, K., Chiles, E., Jankowski, C., Shen, Y., Chen, L., Descamps, H. C., Mukherjee, S., Bhat, Y. R., Lingala, S. R., Chu, Q., Botolin, P., Hayat, F., Doke, T., Susztak, K., Thaiss, C. A., Lu, W., Migaud, M. E., Su, X., Rabinowitz, J. D., … Baur, J. A. (2021). NAD+ flux is maintained in aged mice despite lower tissue concentrations. Cell systems, S2405-4712(21)00338-0. Advance online publication. https://doi.org/10.1016/j.cels.2021.09.001
[2] Xie, N., Zhang, L., Gao, W., Huang, C., Huber, P. E., Zhou, X., Li, C., Shen, G., & Zou, B. (2020). NAD+ metabolism: pathophysiologic mechanisms and therapeutic potential. Signal transduction and targeted therapy, 5(1), 227. https://doi.org/10.1038/s41392-020-00311-7
[3] Liu, L., Su, X., Quinn, W. J., 3rd, Hui, S., Krukenberg, K., Frederick, D. W., Redpath, P., Zhan, L., Chellappa, K., White, E., Migaud, M., Mitchison, T. J., Baur, J. A., & Rabinowitz, J. D. (2018). Quantitative Analysis of NAD Synthesis-Breakdown Fluxes. Cell metabolism, 27(5), 1067–1080.e5. https://doi.org/10.1016/j.cmet.2018.03.018
[4] Schultz, M. B., & Sinclair, D. A. (2016). Why NAD(+) Declines during Aging: It's Destroyed. Cell metabolism, 23(6), 965–966. https://doi.org/10.1016/j.cmet.2016.05.022
[5] Covarrubias, A. J., Perrone, R., Grozio, A., & Verdin, E. (2021). NAD+ metabolism and its roles in cellular processes during ageing. Nature reviews. Molecular cell biology, 22(2), 119–141. https://doi.org/10.1038/s41580-020-00313-x
[6] Mattison, J. A., Colman, R. J., Beasley, T. M., Allison, D. B., Kemnitz, J. W., Roth, G. S., Ingram, D. K., Weindruch, R., de Cabo, R., & Anderson, R. M. (2017). Caloric restriction improves health and survival of rhesus monkeys. Nature communications, 8, 14063. https://doi.org/10.1038/ncomms14063
[7] Song, J., Ke, S. F., Zhou, C. C., Zhang, S. L., Guan, Y. F., Xu, T. Y., Sheng, C. Q., Wang, P., & Miao, C. Y. (2014). Nicotinamide phosphoribosyltransferase is required for the calorie restriction-mediated improvements in oxidative stress, mitochondrial biogenesis, and metabolic adaptation. The journals of gerontology. Series A, Biological sciences and medical sciences, 69(1), 44–57. https://doi.org/10.1093/gerona/glt122
[8] Chalkiadaki, A., & Guarente, L. (2012). Sirtuins mediate mammalian metabolic responses to nutrient availability. Nature reviews. Endocrinology, 8(5), 287–296. https://doi.org/10.1038/nrendo.2011.225
[9] Spindler S. R. (2010). Caloric restriction: from soup to nuts. Ageing research reviews, 9(3), 324–353. https://doi.org/10.1016/j.arr.2009.10.003
[10] Frederick, D. W., Davis, J. G., Dávila, A., Jr, Agarwal, B., Michan, S., Puchowicz, M. A., Nakamaru-Ogiso, E., & Baur, J. A. (2015). Increasing NAD synthesis in muscle via nicotinamide phosphoribosyltransferase is not sufficient to promote oxidative metabolism. The Journal of biological chemistry, 290(3), 1546–1558. https://doi.org/10.1074/jbc.M114.579565
