
Catena | 禁牧后温带草地土壤碳氮磷储量的耦合与解耦(长文翻译)

[题目]
Coupling and decoupling of soil carbon, nitrogen and phosphorus stocks following grazing exclusion in temperate grasslands
[作者]
JiweiLi a b c, Kaibo Wang d, Zhouping Shangguan a b c, Lei Deng c
[单位]
aState Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling, Shaanxi 712100, China
bUniversity of Chinese Academy of Sciences, Beijing 100049, China
cInstitute of Soil and Water Conservation, Northwest A&F University, Yangling, Shaanxi 712100, China
dState Key Laboratory of Loess and Quaternary Geology, Institute of Earth Environment, Chinese Academy of Sciences. Xi’an, Shaanxi, 710061, China
Highlights
(1) Soil C and N stocks significantly accumulated following grazing exclusion.
禁牧后土壤碳氮储量显著增加。
(2) Soil P stocks tended to decrease after grazed exclusion in temperate grassland.
温带草地在禁牧后土壤磷储量呈下降趋势。
(3) SOCand TN stocks were decoupled with TP stocks since grazed exclusion.
由于禁牧,SOC和TN库存与TP库存解耦。
(4) Soil extracellular enzymes played critical roles in enhancing soil N stocks.
土壤胞外酶在提高土壤氮储量中起着重要作用。
Abstract
Grazing exclusion is considered a crucial measure for improving ecosystem structure and function, including elements such as soil carbon and nutrients. 禁止放牧被认为是改善生态系统结构和功能的关键措施,包括土壤碳和养分等元素。
However, little is known about the relationships of soil organic C (SOC), total nitrogen (TN), and total phosphorous (TP) stocks during long-term grazing exclusion. 但长期禁牧期间土壤有机碳(SOC)、总氮(TN)和总磷(TP)储量之间的关系尚不清楚。
This study examined the SOC, TN, and TP stocks after 0–33years of grazing exclusion in temperate grasslands on the Loess Plateau, China. 研究了黄土高原温带草地禁牧0–33年后土壤有机碳、全氮和总磷储量的变化。
The results showed that long-term grazing exclusion significantly increased SOC and TN stocks but declined TP stocks compared to those in grazing grasslands. 结果表明与放牧草地相比,长期禁牧显著提高了土壤有机碳和全氮储量,降低了TP储量;
In addition, grazing exclusion was more favorable for SOC and TN accumulations than TP accretion, and SOC and TN stocks were decoupled from TP stock. 此外,不放牧对土壤有机碳和全氮的积累比TP的积累更有利,且土壤有机碳和全氮的蓄积量与TP的蓄积量呈脱钩关系。
Overall, changes in SOC stock were determined by plant productivity, and TN stocks were also driven by soil microbial activities (i.e., N-acquiring enzymes). 总体而言,土壤有机碳储量的变化受植物生产力的影响,而全氮储量的变化也受土壤微生物活性(即氮获取酶)的影响。
Nevertheless, plant and microbial characteristics had no effects on TP stocks following grazing exclusion. 然而,植物和微生物特征对放牧后全磷储量没有影响。
This study emphasizes the positive effect of grazing exclusion on SOC and TN accumulations, further, it is needed to explore soil phosphorus stabilization mechanisms during grassland restoration after grazing exclusion. 本研究强调了禁牧对土壤有机碳和全氮积累的积极作用,并进一步探讨禁牧后草地恢复过程中土壤磷的稳定机制。
Keywords
Carbon sequestration(碳固定);Grazing exclusion;Nitrogen sequestration;Temperate grassland;Vegetation restoration
1.Introduction
Long-term overloading and improper grazing can destroy thegrassland ecosystem function, which causing severe soil erosion, subsurface compaction and degradation in grasslands around the world. Such degradation can gradually impact the quality of animal husbandry and ecological sustainability, which includes reductions in soil microbial activity and resource availability. Consequently, restoration and governance on degraded grasslands are urgently needed, as they have great capacity to preserve a considerable fraction of atmospheric carbon dioxide under appropriate grazing practices. Grazing exclusion is considered a highly effective measure for the restoration of overgrazed grasslands, by improving plant productivity and soil quality. Therefore, obtaining a better understanding of how grazing exclusion influences soil quality is critical for maintaining grassland functionality and sustainability.长期的超载和不当放牧会破坏草地生态系统功能,造成世界各地草地严重的水土流失、地下压实和退化。这种退化会逐渐影响畜牧业的质量和生态可持续性,包括土壤微生物活性和资源可用性的减少。因此,退化草地的恢复和治理是迫切需要的,因为在适当的放牧做法下,它们有很大的能力保存大气中相当一部分的二氧化碳。禁牧被认为是通过提高植物生产力和土壤质量来恢复过度放牧草地的一种非常有效的措施。因此,更好地理解禁牧如何影响土壤质量对于保持草地功能和可持续性至关重要。
Soil organic carbon (SOC), total nitrogen (TN), and phosphorus (TP) stocks, and cycling processes are important aspects of sustainable soil quality, and the ecosystem functions of grasslands. Grazing exclusion can directly affect the distribution of SOC, TN, and TP, and exert complex effects onsoil organic matter (SOM) cycling. However, the responses of SOC, TN, and TP stocks to grazing exclusion have been controversial in grasslands. Some studies have revealed that grazing exclusion leads to increased SOC and nutrients, whereas others have shown that there were negative or even no significant impacts of grazing exclusion on SOC, TN, and TP immobilization in grasslands. These inconsistencies are likely due to variations in grassland types, enclosure durations, grazing history, and factors limiting plant growth; thereby affecting ecosystem SOM cycling in these regions. Thus, the effects of grazing exclusion periods on SOC and nutrients in grasslands remain unclear. Therefore, we aimed to clarify the response and drivers of SOC, TN, and TP stocks and to scientifically explore a grassland management strategy for improving the production and ecological functions of grasslands through grazing exclusion. 土壤有机碳(SOC)、全氮(TN)和全磷(TP))储量及其循环过程是影响草地土壤可持续质量和生态系统功能的重要方面。禁牧直接影响土壤有机碳、全氮和全磷的分布,并对土壤有机质循环产生复杂的影响。然而,草地SOC、TN和TP储量对禁牧的响应一直存在争议。一些研究表明,禁牧导致草地SOC和养分增加,而另一些研究表明,禁牧对草地SOC、TN和TP固定具有负效应甚至不显著的影响。这些不一致性可能是由于草地类型、封育时间、放牧历史和限制植物生长的因素的差异;从而影响这些区域的生态系统有机质循环。因此,禁牧期对草地有机碳和养分的影响尚不清楚。因此,本研究旨在阐明SOC、TN和TP储量的响应及其驱动因素,科学探索通过禁牧提高草地生产和生态功能的草地管理策略。
Ecological restoration via grazing exclusion can alter the distributions of SOC, TN, and TP in grassland ecosystems. The persistence and stabilization of SOC, TN, and TP stocks are altered by biotic and abiotic factors, including vegetation characteristics, aggregate physical properties, and microbial nutrient use efficiency. In general, greater plant productivity and microbial biomass represent greater root surface areas and higher SOC and TN stocks. Previous studies have focused on the impact of biological and abiotic factors, such as grazing exclusion period, soil physicochemical variables, plant characteristics, and microorganismson the variations in TN and SOC sequestration in grassland ecosystems. Nevertheless, most of these studies have only compared single un-grazed grasslands against grazed grasslands and focused on the single periods of exclusion. In addition, these studies have mostly ignored the combined impacts of biotic and abiotic variables on SOC, TN, and TP stocks. Therefore, we aimed to clarify the proportionate contributions of microbial, plant, and edaphic variables on TN, TP, and SOC stocks; and to scientifically explore a grassland management strategy for improving the ecological functions of grasslands through grazing exclusion practices. 通过放牧进行生态恢复可以改变草地生态系统SOC、TN和TP的分布。植被特征、团聚体物理性质和微生物养分利用效率等生物和非生物因素影响着土壤SOC、TN和TP储量的持久性和稳定性。总体而言,较高的植物生产力和微生物生物量代表较大的根系表面积和较高的SOC和TN储量。以往的研究主要集中在放牧封育期、土壤理化变量、植物特征和微生物等生物和非生物因素对草地生态系统全氮和有机碳固存变化的影响。然而,这些研究大多只比较了单一的未放牧草地和放牧草地,并集中在单一的排除时期。此外,这些研究大多忽略了生物和非生物变量对SOC、TN和TP储量的联合影响。因此,本研究旨在阐明微生物、植物和土壤变量对全氮、全磷和有机碳储量的比例贡献;科学探索通过禁牧改善草地生态功能的草地管理策略。
Grazing exclusion facilitatesSOM formation and stability by altering both the quality and quantity of litter and root input, and consequently, indirectly shifts in SOC, TN, and TP dynamics. To our knowledge, most studies have principally focused on comparing SOC stocks of grazed and un-grazed lands; however, there are limited studies on nutrient stocks (especially TP stocks). In addition, soil TP stocks have been mainly derived from rock weathering, whereas soil TN stocks are influenced by nitrogen fixation, deposition input, and inorganic nitrogen leaching. Previous studies have proposed that soil TN and TP dynamics may strictly limit the soil carbon (C) accumulation during vegetation recolonization in natural terrestrial ecosystems. In general, SOC accumulation is coupled to TN because of its influence on SOM decomposition. However, experiments aimed at investigating changes in soil TN and TP stocks have not been attempted, and few studies have paid close attention to the associations between SOC, TN, and TP stocks following grazing exclusion. Therefore, it is necessary to evaluate the responses of C and nutrient stocks to grazing exclusion. 禁牧通过改变凋落物和根系输入的质量和数量,促进SOM的形成和稳定,从而间接改变SOC、TN和TP的动态。据我们所知,大多数研究主要集中在比较放牧和未放牧土地的有机碳储量;然而,对营养储量(特别是总磷储量)的研究有限。此外,土壤TP储量主要来源于岩石风化,而土壤TN储量受氮固定、沉积输入和无机氮淋溶的影响。已有研究表明,在自然陆地生态系统中,土壤TN和TP动态可能严格限制了植被再定植过程中土壤碳(C)的积累。总体上,SOC的积累与TN的耦合作用影响SOM的分解。然而,目前还没有针对土壤全氮和全磷储量变化的试验,而且很少有研究关注放牧后土壤有机碳、全氮和全磷储量之间的关系。因此,有必要评估碳储量和养分储量对禁牧的响应。
To recover ecosystem function in theLoess Plateau region, the Chinese government has implemented the ‘Returning Grazing Land to Grassland’ project, which prohibits grazing in many grasslands through grazing exclusion. Vegetation characteristics, soil physical and chemical properties, and microbial activities may be altered through grazing exclusion. However, there is limited information on the effects of long-term grazing exclusion strategies on SOC, TN, and TP stocks. In addition, limited information is available about which microbial, plant, and edaphic variables have the impacts on the stocks of SOC, TN, and TP. To address these knowledge gaps, this study sampled soils that represent a natural successional sequence following grazing exclusion for 0, 8, 18, 26, and 33 years on the Chinese Loess Plateau; to (1) examine how SOC, TN, and TP stocks responded, and (2) to identify the key microbial, plant, and edaphic variables that affect SOC, TN, and TP stocks after grazing exclusion. 为了恢复黄土高原地区的生态系统功能,中国政府实施了“退牧还草”工程,在许多草原上实行禁牧。植被特征、土壤理化性质和微生物活动可能会通过放牧被改变。然而,目前关于长期禁牧策略对SOC、TN和TP储量的影响的信息有限。此外,关于哪些微生物、植物和土壤变量对SOC、TN和TP储量有影响的信息有限。为了填补这些知识空白,本研究以黄土高原放牧0、8、18、26和33年的自然演替序列为样本;(1)研究SOC、TN和TP储量的响应;(2)识别放牧后影响SOC、TN和TP储量的关键微生物、植物和土壤变量。
2.Materials and methods
2.1.Study area and field sampling
The study area was the Yunwu Mountain grassland, Ningxia Province, China, located northeast of Guyuan and in the middle of theLoess Plateau (36°10′–36°17′N, 109°16′–109°24′E; altitude, 1700–2148 m above sea level). Since 1980, the government has implemented several strict measures to exclude livestock and farmers for ecological restoration purposes in the Yunwu Mountain region. The area thus provides a range of exclusion ages for study. The annual average temperature is 7 °C, and the annual average precipitation is 425 mm, 60%–75% of the precipitation occurs in summer between July and September, and atmospheric precipitation in the main water supply. The area has a typical temperate semi-arid climate, and the vegetation is typical of temperate grassland. The soil type in the study area was aeolian soil (silt loam), and the dominant plant species were Stipa grandis, Potentilla acaulis, Artemisia capillaris, and Thymus mongolicus.
In the experimental area, achronosequence of five grasslands with different grazing exclusion periods (0, 8, 18, 26, and 33 years) was selected to investigate when the aboveground biomass peaked based on measurements in 2019. Grazed grassland was used as a control (GE0), consisting of an unfenced free-grazing community. All sites had similar soil types and geographical characteristics (Table S1). At each site, six plots (20 m × 20 m) were established at least 10 m apart as independent replicates. Within each plot, three quadrats (1 m × 1 m) were randomly established, and in each quadrat, plant community coverage (PCC), plant community height (PCH), litter biomass (LB), belowground biomass (BB), and aboveground biomass (AB) were measured as described by Deng et al. (2014). For each of the five grassland ages, twenty upper mineral soil cores (0–10 cm) were randomly collected from each layer in each plot and mixed thoroughly to produce a composite sample using a 2-mm sieve after removing the aboveground tissues. Then, the soil samples were placed in a portable cooler box and transported to the laboratory for further measurement. The soil samples were subsequently divided into two parts: one part was applied to measure soil extracellular enzyme activities and microbial biomass at 4 °C for up to 2 weeks, and the other part was used to text soil physical and chemistry properties with air-drying at 25 °C.
2.2.Soil physical and chemistry properties
Soil bulk density (BD, g cm−3) were determined by the cutting ring method.Soil organic carbon (SOC) and total nitrogen(TN) were determined using the calorific boiling and Kjeldahl method, respectively (Nelson and Sommers, 1983). A suspension in a 1: 2.5:1 ratio of soil to water was applied to measure soil pH by a pH meter (Mettler Toledo, USA). Soil moisture content (SW, %) was determined gravimetrically by weight loss after oven-drying 100 g of fresh soil at 105 °C for 48 h. Soil particle size distribution was analyzed using a Mastersizer 2000 laser sizer (Malvern Instruments Ltd., Malvern, UK), and was, then, divided into sand, silt, and clay. Dissolved organic carbon (DOC) concentrations were measured using a total organic carbon (TOC) analyzer. Soil available N(SAN) concentration were analysed by the continuous alkali-hydrolyzed reduction diffusing (Stanford, 1982). The contents of soil available phosphorus(SAP) and total phosphorus(TP) were measured by an ultraviolet spectrophotometer (Hitachi UV2300) after extracted with 0.5 M NaHCO3 and H2SO4-HClO4, respectively.
2.3. Soil extracellular enzyme activities and microbial biomass
The activities of threehydrolytic enzymes (β-1,4-glucosidase, BG; β-1,4-N-acetylglucosaminidase, NAG; alkaline phosphatase, ALP) were assayed. The activities of BG for microbial C-acquisition, NAG for microbial N-acquisition, and ALP for microbial P-acquiring. Fresh soil (eight duplicate) was dispensed into 96-well microplates with buffer solution. Fluorescence were assayed by a microplate reader with 450-nm emission and 365-nm excitation filte after the solution incubated at 25 °C for 4 h in the dark. The activities of soil extracellular enzyme were expressed as µmol g−1 h−1. The chloroform fumigation extraction method was used to determine the soil microbial biomass carbon (MBC) and nitrogen (MBN), extracting using K2SO4 (100 mL 0.5 mol L-1) (Brookes et al., 1985). The extractions were measured for MBC and MBN using a Total Organic C/N analyzer. Conversion factors used to calculate MBC and MBN were 0.45 and 0.54 by Brookes et al. (1985), respectively. Microbial biomass phosphorus (MBP) was extracted by 0.5 M NaHCO3, using the fumigation extraction method described by Brookes et al. (1982). The experimental conversion factor for calculating MBP was 0.40 (Brookes et al., 1982).
2.4.Estimation of the TN, TP, and SOC stocks
TN, TP, and SOC stocks were calculate by the following equations (Li et al., 2020a):
whereX denotes the SOC, TN, or TP stocks after grazing exclusion, TN is the soil total N content (g kg−1), BD is the soil bulk density (kg m−3), SOC is the soil organic C content (g kg−1), TP is the total phosphorus content, and D is the soil layer thickness (cm).
2.5.Calculation of TN, TP, and SOC sequestration
We used the following equations to calculate SOC, TN, and TP sequestrations (Deng et al., 2017,Li et al., 2012):
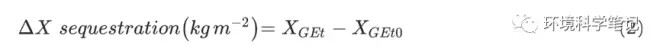
where △X denote the variations in the stocks of TN, TP, or SOC after grazing exclusion, respectively; XGEt0 are the stocks of TN, TP, or SOC in the grazing grassland; and XGEt express stocks of SOC, TN, or TP at the site after grazing exclusion for t years, respectively.
2.6.Estimation of the change rates of SOC, TN, and TPstocks
We applied the following equations to estimate the rates of relative change in the stocks of TN, TP, or SOC (Deng et al., 2016,Li et al., 2012, Li et al., 2020a):
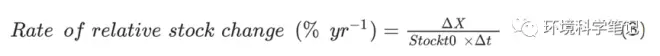
where stockt0is the TN, TP, or SOC stocks in the grazing grassland, Δt denotes the time (years) since after grazing exclusion, and ΔX express the change in the stocks of TN, TP, or SOC after grazing exclusion.
2.7.Statistical analysis
Linear mixed-effects models (LMM) were used to test the effects of grazing exclusion time, soil depth, and their interaction on each variable using the ‘nlme’ function in R (version 4.0). When analyzing the SOC, TN, or TP stocks andsoil physicochemical properties, ‘soil depth’, ‘grazing exclusion’, and their ‘interaction’ were used as the fixed factors, whereas ‘plot’ was used as a random factor. Redundancy analysis (RDA) was also used to determine the effects of plant and soil variables on the SOC, TN, and TP stocks with the ‘vegan’ package in R (version 4.0). To elucidate the relative influences of microbial, edaphic and vegetation variables on the SOC, TN, and TP stocks, multivariable linear regression analysis (MLRA) was performed the relative importance of plant, soil, and microbial variables to explain the SOC, TN, or TP stocks. Before the analysis, variables with high variance inflation factors (VIF > 20) were eliminated from the model to exclude collinearity. However, no factor was found to control TP stocks (R2 = 0.11, P = 0.40). Structural equation modeling (SEM) frameworks were then applied to reveal both the indirect and direct impacts of the driving variables on the TN, TP, and SOC stocks in AMOS 23.0.
3.Results
3.1.Variations in TN, TP, and SOC stocks since grazing exclusion
Compared with stocks in the free-grazing site, grazing exclusion significantly increased SOC stocks (SOCS) and TN stocks (TNS) but decreased TP stocks (TPS,P < 0.05). In the 0–20 cm layer, 26 and 33 years of grazing exclusion significantly increased SOCS by 216.8% and 207.0%, respectively (Fig. 1a); whereas in the initial 18 years after grazing exclusion, no significant differences in SOCS were found in the 40–60 cm layer (Fig. 1a). After 33 years of grazing exclusion, TNS was significantly higher than in the initial 26 years following grazing exclusion in the 0–20 cm layer (Fig. 1b), but there was little difference in the TNS after grazing exclusion in the 40–60 cm layers. In contrast, grazing exclusion resulted in a significantly lower TPS in the 0–40 cm soil layer (P < 0.05) (Fig. 1c) than in free-grazing sites.
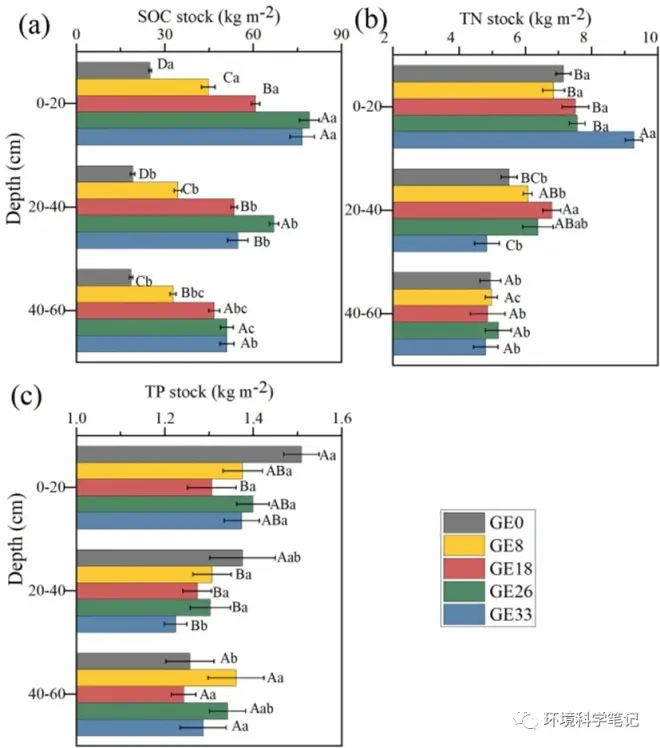
Fig. 1.Effects of grazing exclusion on the distribution of soil organic carbon (SOC, a), total nitrogen (TN, b), and total phosphorus (TP, c) stocks at 0–60 soil layer.
The values of ΔSOCS at the depths of 0–60cm significantly increased after 8 years grazing exclusion, but ΔSOCS presented no significant differences after 26 and 18 years of grazing exclusion in the 0–60 cm layers (Fig. 2a). Grazing exclusion for 33 years significantly increased ΔTNS in the 0–20 cm layer, but significantly decreased ΔTNS in the 20–40 cm layer (Fig. 2b). Soil TPS showed no significant differences after grazing exclusion (Fig. 1c). The rates of relative changes in SOCS and TNS decreased continuously across the 33-year period of grazing exclusion (Fig. 2d and e). The values of ΔTNS and ΔTPS showed no significant changes in the 0–60 cm layer (Fig. 2e and f).
Fig. 2. Effects of grazing exclusion on the sequestrations (a-c) and accumulation rates (d-f) of soil organic carbon (SOC, a), total nitrogen (TN, b), and total phosphorus (TP, c) stocks at 0–60 soil layer.
3.2.Relationships of soil TN, TP, and SOC changes since grazing exclusion
The relationships among SOC, TN, and TP pools exhibited diverse trends after grazing exclusion (Fig. 3). Significant relationships were found between the shifts in the SOC and TN contents (R2= 0.44, P < 0.001, y = 8.53x-1.62; Fig. 2a), SOC and TP contents (R2 = 0.38, P < 0.001, y = 90.86x-29.37; Fig. 2b), and TN and TP contents (R2 = 0.20, P < 0.001, y = 5.01x-0.21; Fig. 2c). However, there were no significant correlations between the changes in the stocks and sequestrations of SOC, TN, and TP, except for a highly correlated relationship between changes in SOCS and TNS (R2 = 0.17, P < 0.001, y = 5.35x + 14.58; Fig. 2d-i).
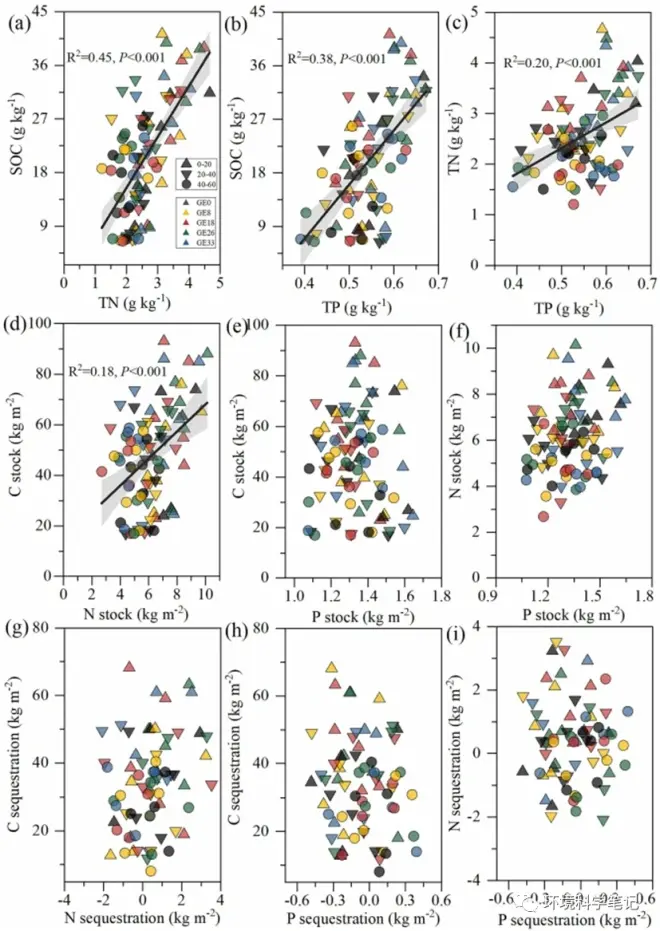
Fig. 3.Regression relationships of SOC, TN, and TP in contents, stocks, and sequestrations after grazing exclusion. Gray lines indicate 95% confidence intervals, while black lines are the linear model fits of these linear models between soil SOC and TN, TP in sequestrations and accumulation rates. Note: SOC, soil organic carbon; TN, total nitrogen; TP, total phosphorus.
3.3.Factors correlated with the TN, TP, and SOC stocks
The RDA analysis revealed that the RDA1 and RDA2 axes explained 95.35% and 4.59%, respectively, of the variation in the SOCS, TNS, and TPS in the 0–60cm layer (Fig. 4). SOCS was positively correlated with most of the vegetation characteristics, including PCC, PCH, AB, LB, and MBC. In contrast, the TPS were not associated with any environmental factors (Fig. 5c). The results of the MLRM showed that the environmental factors explained 82.1% of the variability in SOCS (Fig. 5a) and 58.9% of the variability in TNS (Fig. 5b). The variations in SOCS were best explained by PCC, LB and DOC (Fig. 5a), whereas NAG, AB, and BB had strong influences on the variation in TNS (Fig. 5b). Furthermore, none of the measured factors were found to control TPS.
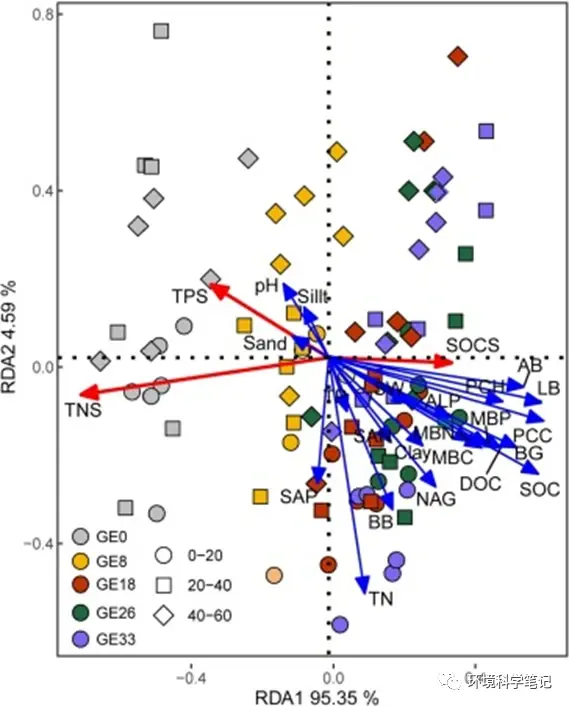
Fig. 4.Redundancy analysis (RDA) ordination diagram for SOC, TN, and TP stocks with environmental variables at 0–60 cm soil layer as affected by grazing exclusion. Note: SOC, soil organic carbon; TN, total nitrogen; TP, total phosphorus; AB, aboveground biomass; BB, belowground biomass; PCC, plant community coverage; LB, litter biomass; PCH, plant community height; pH, soil pH; BD, bulk density; SW, soil water; DOC, dissolved organic carbon; SAN, available nitrogen; SAP, available phosphorus; MBC, microbial biomass carbon; MBN, microbial biomass nitrogen; MBP, microbial biomass phosphorous; BG, β-1,4-glucosidase; NAG, β-1,4-N-acetylglucosaminidase; ALP, alkaline phosphatase.
Fig. 5.Effects of plant, soil, and microbial properties on SOC (a) and TN (b) stocks from multivariable linear regression model and the Pearson correlation matrix analysis among SOC, TN, and TP stocks and environmental variables at 0–60 cm soil layer as affected by grazing exclusion (c). No factor was found to control TP stock (R2 = 0.11, P = 0.40). Note: Red indicates positive correlations, and blue indicates negative with environmental variables in Pearson correlation matrix analysis. SOCS, soil organic carbon stock; TNS, total nitrogen stock; TPS, total phosphorus stock; SOC, soil organic carbon; TN, total nitrogen; TP, total phosphorus; AB, aboveground biomass; BB, belowground biomass; PCC, plant community coverage; LB, litter biomass; PCH, plant community height; pH, soil pH; BD, bulk density; SW, soil water; DOC, dissolved organic carbon; SAN, available nitrogen; SAP, available phosphorus; MBC, microbial biomass carbon; MBN, microbial biomass nitrogen; MBP, microbial biomass phosphorous; BG, β-1,4-glucosidase; NAG, β-1,4-N-acetylglucosaminidase; ALP, alkaline phosphatase.
The SEM models showed that the critical factors explained 75% and 57% of the variation in the SOCS (Fig. 6a) and TNS (Fig. 6b), respectively. Grazing exclusion significantly affected the effect of PCC and LB on SOCS variation (Fig. 6a), and LB and DOC directly affected SOCS (Fig. 6a). Moreover, grazing exclusion exerted significant positive impacts on BB and NAG, but a negative effect on AB in affecting TNS variation (Fig. 6b). BB and NAG showed remarkably favorable impacts on the TNS (Fig. 6b), whereas it had a significantly negative and direct impact on the TNS variation. Additionally, the total standardized effects of grazing exclusion, PCC, LB, and DOC on SOCS were 0.82, 0.45, 0.36, and 0.34, respectively (Fig. 6c), and the effects of grazing exclusion, BB, AB and NAG on TNS were 0.13, 0.67, −0.42 and 0.48, respectively (Fig. 6d).
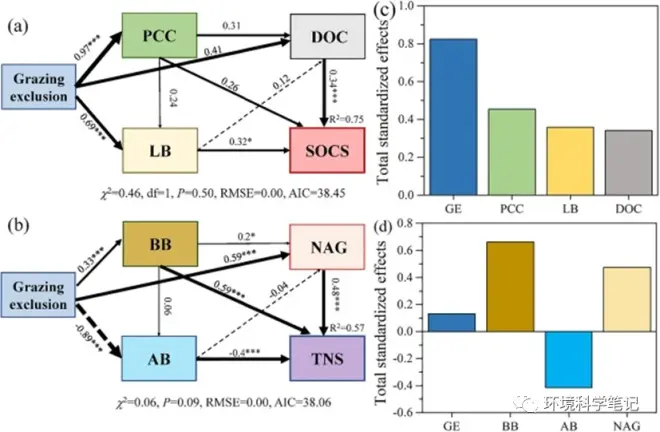
Fig. 6.The structural equation model (SEM) examining the effects of key factors on the contributions of the direct and indirect regulatory pathways of SOC and TN stocks at 0–60 soil layer (no factor was found to control TP stock). Note: SOCS, soil organic carbon stock; TNS, total nitrogen stock; TP, total phosphorus; AB, aboveground biomass; BB, belowground biomass; PCC, plant community coverage; LB, litter biomass; PCH, plant community height; pH, soil pH; BD, bulk density; SW, soil water; DOC, dissolved organic carbon; SAN, available nitrogen; SAP, available phosphorus; MBC, microbial biomass carbon; MBN, microbial biomass nitrogen; MBP, microbial biomass phosphorous; BG, β-1,4-glucosidase; NAG, β-1,4-N-acetylglucosaminidase; ALP, alkaline phosphatase.
4.Discussion
4.1.Effects of grazing exclusion on SOC, TN, and TP stocks
Changes in the accumulation of SOC, TN, and TP following grazing exclusion play critical roles in estimating land recovery in degraded grasslands, and forecasting the effects of grassland recovery on global climate warming. However, the dynamics of SOC, TN, and TP cycles following grazing exclusion are rather complex, and few studies have focused on TP accumulation in arid and semi-arid regions. Based on our results, grazing exclusion (8–33years) significantly increased SOC stock and sequestration in the 0–60 cm soil profile (Fig. 1a). The increases observed in our study may have been caused by the increased inputs of plant and litter biomass after the grazing exclusion (Table S1). Generally, grazing exclusion reduces soil compaction by livestock trampling and improves soil water holding capacity and plant productivity, thereby leading to higher soil C stocks. However, some studies have reported that grazing exclusion has a negative effect on SOC accumulation. This inconsistency may be due to the shorter recovery periods, low precipitation, and extreme cold conditions in the study areas, which limit vegetation development. 禁牧后土壤有机碳(SOC)、总氮(TN)和总磷(TP)积累的变化对评估退化草地土地恢复、预测草地恢复对全球气候变暖的影响具有重要意义。然而,禁牧后土壤有机碳、全氮和总磷的动态变化较为复杂,对干旱半干旱区土壤总磷积累的研究较少。根据我们的结果,不放牧(8-33年)显著增加了0-60 cm土壤剖面上的有机碳存量和固相量(图1a)。在我们的研究中观察到的增加可能是由于排除放牧后植物和凋落物生物量投入的增加造成的(表S1)。一般来说,禁牧减少了家畜踩踏造成的土壤压实,提高了土壤持水能力和植物生产力,从而提高了土壤碳储量。但也有研究表明,禁牧对土壤有机碳积累有负面影响。这种不一致可能是由于研究地区的恢复周期较短、降水较少和极端寒冷的条件限制了植被的生长。
Compared to those on SOCS, there are relatively few reports on soil TNS and TPS following grazing exclusion. In our study, grazing exclusion decreased TP stock (Fig. 1c) and sequestration (Fig. 2c), whereas grazing exclusion for 33 years significantly increased TN stock (Fig. 1b) and sequestration (Fig. 2b) within the 0–20 cm soil layer. Generally, terrestrial ecosystem nitrogen (N) pools rely on atmospheric N deposition, plant N fixation or both. The high soil TNS in the grazing exclusion sites was mainly linked to plant litter return. Importantly, 33 years of grazing exclusion can provide sufficient N stocks, which may be related to the N-fixing species plant species that fix N from the atmosphere. In addition, the declines in soil TPS due to grazing exclusion may be related to soil leaching and plant consumption. As the new phosphorus (P) is mainly derived from rock weathering, grazing exclusion leads to high soil P loss through the promotion of plant cover and soil moisture recovery. Our results showed a significant correlation between soil P content and soil water (Fig. 5c), suggesting that soil P loss is to some extent linked to water erosion. Furthermore, soil TNS and TPS showed no significant differences in the 20–60 cm layer with grazing exclusion (Fig. 1b and c), indicating that deeper soils such as the 20–60 cm layer are less sensitive to grazing exclusion than topsoil layers, and potentially, that extended restoration periods are needed for noticeable accumulation of soil TNS and TPS to occur in the subsoil. 与SOCS相比,禁牧后土壤TNS和TPS的研究报道相对较少。在我们的研究中,禁牧降低了全氮储量(图1c)和固存量(图2c),而33年的禁牧显著增加了0-20 cm土层的全氮储量(图1b)和固存量(图2b)。通常,陆地生态系统氮库依赖于大气氮沉降、植物氮固定或两者兼有。禁牧地高土壤TNS主要与植物凋落物归还有关。重要的是,33年的禁牧可以提供足够的氮储量,这可能与固定氮的植物物种从大气中固定氮有关。此外,由于禁牧导致的土壤TPS下降可能与土壤淋洗和植物消耗有关。由于新磷主要来源于岩石风化,禁牧通过促进植物覆盖和土壤水分恢复导致土壤磷流失。我们的结果表明,土壤磷含量与土壤水分之间存在显著的相关性(图5c),表明土壤磷的流失在一定程度上与水土流失有关。此外,在禁牧条件下,20-60 cm土层的土壤总养分和总养分利用率没有显著差异(图1b和c),这表明较深层的土壤(如20-60 cm土层)对禁牧的敏感性低于表层土壤,而且可能需要延长恢复期才能使土壤总养分和总养分利用率显著地积累到下层土壤中。
4.2.Interactions of SOC, TN, and TP accumulations after grazing exclusion
The interactions of SOC, TN, and TP play key roles in terrestrial ecosystemsustainability. Grazing exclusion can promote vegetation succession and recolonization, leading to redistribution of SOC, TN and TP. We found that the contents of SOC, TN, and TP showed a positive correlation following grazing exclusion (Fig. 3a-c). These findings are consistent with those of the previous studies. There are limited results regarding the correlations between soil TP changes and SOC and TN accumulation following grazing exclusion. Interestingly, our results only found a close relationship between SOCS and TNS (Fig. 3d), but not between SOCS and TPS, or TNS and TPS, as well as their rates of accumulation (Fig. 3e-i). These results indicate that SOC, TN, and TP sequestration were decoupled after grazing exclusion. In general, grazing exclusion increases SOC and TN stocks because there are sufficient inputs of plant residue in the grassland ecosystem; and because fresh P is mainly derived from rock weathering, it might not adapt the elevated SOC and TN in soil. As the SAP from soil resources has been depleted, plants and soil microorganisms may acquire more P to meet their growth. What 's more, resource P deficiency might aggravate the competition between vegetation and soil microorganisms under high soil moisture levels following grazing exclusion. Therefore, soil TPS would be decoupled from soil C and N stocks. SOC、TN和TP的交互作用对陆地生态系统的可持续性具有关键作用。禁牧可促进植被演替和再定植,导致SOC、TN和TP的再分布。结果表明:禁牧后,土壤SOC、TN、TP含量呈显著正相关(图3a-c)。这些发现与之前的研究结果一致。目前关于禁牧后土壤TP变化与SOC和TN积累的相关性研究结果有限。有趣的是,我们的结果只发现了SOCS和TNS之间的密切关系(图3d),而没有发现SOCS和TPS之间,或者TNS和TPS之间,以及它们的累积速率之间的密切关系(图3e-i)。综上所述,禁牧后,土壤SOC、TN和TP固存量均出现去耦。总体而言,禁牧增加了草地生态系统中充足的植物残渣输入,增加了SOC和TN储量;新鲜P主要来源于岩石风化,可能不适应土壤SOC和TN的升高。随着土壤资源的枯竭,植物和土壤微生物可能会获得更多的磷以满足其生长。此外,资源磷缺乏可能会加剧放牧后高水分条件下植被与土壤微生物之间的竞争。因此,土壤TPS将与土壤碳和氮储量脱钩。
4.3.Factors controlling SOC, TN, and TP stocks after grazing exclusion
Grazing exclusion increased vegetation cover, plant community height, and plant biomass as a result of decreased livestock trampling and BD. Thus, soil microorganisms may increase their investments in extracellular enzymes in response to increases in litter andrhizosphere inputs after grazing exclusion. Our results revealed that SOCS was correlated with most of the measured plant, soil, and microbial variables (Fig. 5a), suggesting that it is sensitive to the environmental changes following grazing exclusion. Among them, plant cover had the greatest effect on SOCS (Fig. 5a and Fig. 6c), indicating that vegetation community characteristics play critical roles in soil C accumulation. First, grazing exclusion reduces consumption and trampling of plants by livestock and promotes the recovery of the soil microenvironment (e.g., soil temperature, BD, and moisture) and vegetation structure. Second, changes in the microenvironment facilitate the decomposition of the continuous input of litter from various types of vegetation, which acts as a major source of soil C accumulation. It has been demonstrated that after vegetation cover, LB is the second most important factor affecting soil C accumulation (Fig. 5a and Fig. 6c). Third, surface litter and humus decomposition can alter the community structure and activities of soil microbes, contributing to the formation of macroaggregates. Soil aggregates can protect SOM through a wide variety of pathways, from incorporation to adsorption, resulting in SOC accumulation. Finally, increments in the vegetation cover can alleviate SOC loss by hydraulic and wind erosion. The complex impacts of these factors result in increases in SOCS and are accompanied by extensions in vegetation cover. 禁牧增加了植被盖度、植物群落高度和植物生物量。因此,土壤微生物可能增加了对细胞外酶的投资,以响应禁牧后枯落物和根际输入的增加。我们的结果表明,SOCS与大部分测量的植物、土壤和微生物变量相关(图5a),表明它对禁牧后的环境变化很敏感。其中,植物覆盖度对SOCS的影响最大(图5a和图6c),说明植被群落特征对土壤碳积累起关键作用。首先,禁牧减少了牲畜对植物的消耗和践踏,促进了土壤微环境(如土壤温度、BD和水分)和植被结构的恢复。其次,微环境的变化促进了各类植被连续输入的凋落物的分解,是土壤碳积累的主要来源。研究表明,在植被覆盖之后,LB是影响土壤C积累的第二重要因素(图5a和图6c)。第三,地表枯落物和腐殖质分解可以改变土壤微生物的群落结构和活性,促进大团聚体的形成。土壤团聚体可以通过多种途径保护SOM,从并入到吸附,导致SOC积累。最后,植被覆盖度的增加可以缓解水蚀和风蚀造成的SOC损失。这些因素的复杂影响导致了SOCS的增加,并伴随着植被覆盖的扩展。
In contrast to SOCS, belowground biomass and NAG exhibited strong impacts on TNS (Fig. 5b), indicating that plant productivity and soil extracellular enzymes play critical roles in TN accumulation following grazing exclusion. This is likely linked to the lack of atmospheric N inputs, because the atmospheric atmosphere N acquired by plants is shifted to the soil. Furthermore, a greater increase in soil C than in N in the 33-year succession likely caused the soil C:N ratio to increase (Fig. S1a), which likely increased soil N limitation. Grazing exclusion allows the plant diversity and vegetation community structures to recover; therefore, successive increases in plant productivity can ensure sufficient resources for microbial mineralization and N fixation. Thus, soil N accumulation may depend on the decomposition of roots and mycorrhiza by soil extracellular enzymes (especially N-acquiring enzymes) in soil microbial communities under N-limited consitions. As such, differences in soil microbial species appear to be strong determinants of soil N accumulation. However, our results showed that soil microbial properties, soil physical and chemistry properties, and vegetation characteristics could not illustrate the main varieties in TPS during grazing exclusion (Fig. 5c). Because of elevated water and nutrient availability (Table S2), this occurrence seems to be counter-intuitive, and the possible cause might be the leaching and exhaustion of TP. In addition, soil erosion also accelerates P loss on the Loess Plateau, and primary minerals become depleted as soil development proceeds, thus leading to decreases in the amount of P released by weathering. Strong competition between vegetation and soil microbial communities is also a key factor that leads to P insufficiency during grassland succession. Therefore, further studies are need to investigate the complex interactions between composition of microbial and plant communities with SOC, TN, and TP accumulation to grazing exclusion. 与SOCS相比,地下生物量和NAG对TNS的影响更大(图5b),表明植物生产力和土壤胞外酶在禁牧后的全氮积累中起着关键作用。这可能与缺乏大气氮输入有关,因为植物获得的大气氮会转移到土壤中。此外,在33年的演替过程中,土壤C的增幅超过N,这可能导致土壤C:N比值增加(图S1a),这可能增加了土壤N限制。禁牧使植物多样性和植被群落结构得以恢复;因此,连续提高植物生产力可以保证充足的资源用于微生物矿化和氮固定。因此,在氮有限的条件下,土壤氮的积累可能依赖于土壤微生物群落中土壤细胞外酶(尤其是氮获取酶)对根和菌根的分解。因此,土壤微生物种类的差异似乎是土壤氮积累的强决定因素。然而,我们的结果表明,土壤微生物特性、土壤理化特性和植被特征并不能说明放牧封育期间TPS的主要变化(图5c)。由于水分和养分利用率的提高(表S2),这种情况似乎与直觉不符,可能的原因可能是TP的淋溶和耗竭。此外,黄土高原的土壤侵蚀也加速了磷的流失,随着土壤发育,原生矿物质逐渐枯竭,从而导致风化释放的磷量减少。植被与土壤微生物群落之间的激烈竞争也是导致草地演替过程中磷不足的关键因素。因此,需要进一步研究微生物和植物群落组成与SOC、TN和TP积累对禁牧的复杂交互作用。
5.Conclusions
Grazing exclusion significantly increased SOCS at soil depths of 0–60cm, indicating that semiarid grasslands with fencing management can act as sinks for C sequestration. Grazing exclusion significantly enhanced TNS, but decreased TPS in the 0–40 cm soil layer. However, there were no significant differences in the TNS and TPS in the 40–60 cm soil layer during the 33 years following grazing exclusion. Furthermore, grazing exclusion is more favorable for SOC and TN accumulations than TP accretion, indicating that SOCS and TNS were decoupled from TP stock. Grazing exclusion had more effects on TN and TP stocks in the topsoil than in the subsoil. Our results showed that plant community characteristics, such as cover and productivity, played an important role in SOC accumulation. Moreover, soil extracellular enzymes (especially N-acquiring enzymes) in soil microbial communities were found to play a key role in shaping soil N accumulation, whereas no factors were found to control variations in TPS following grazing exclusion. Overall, our findings are crucial for assessing the grazing and fencing measures linked to SOC, TN, and TP storages, and for the optimal recovery of degraded grasslands in semiarid areas. 禁牧显著提高了0 ~ 60 cm土层SOCS,表明围封管理的半干旱草地可作为碳固存的汇。禁牧显著提高了0 ~ 40 cm土层TNS,但降低了TPS。而禁牧后33年间,40 ~ 60 cm土层TNS和TPS均无显著差异。此外,放牧排斥比TP积累更有利于SOC和TN的积累,表明SOCS和TNS与TP存量脱钩。放牧排斥对表层土壤TN和TP储量的影响大于对底层土壤。结果表明:覆盖度、生产力等植物群落特征在土壤有机碳积累中发挥重要作用。此外,研究发现土壤微生物群落中的土壤细胞外酶(尤其是氮获取酶)对土壤氮积累起关键作用,而没有发现放牧后土壤微生物TPS的变化。综上所述,本研究结果对评估半干旱区退化草地的放牧和封育措施与SOC、TN和TP储量的关系,以及对退化草地的最佳恢复具有重要意义。
