
生物化学第五章糖代谢(四)
由非糖化合物(乳酸、甘油、生糖氨基酸)转变为葡萄糖或糖原的过程称为糖异生。

糖异生的主要器官是肝。肾糖异生相对较弱,但长期饥饿时可增强。发生部位是细胞质和线粒体。

反应过程:糖异生途径与酵解途径大多数反应是共有的、可逆的。糖异生不完全是糖酵解的逆反应。(糖酵解的三个限速步骤的逆反应由糖异生特有的关键酶催化)
1)丙酮酸经丙酮酸羧化支路生成磷酸烯醇式丙酮酸。
糖异生时需要丙酮酸进入线粒体,启动丙酮酸羧化支路。
两步反应:第一个反应的关键酶是丙酮酸羧化酶,辅因子是生物素。CO2先于生物素结合,需消耗ATP;然后活化的CO2再转移给丙酮酸生成草酰乙酸。反应在线粒体。
第二个反应的关键酶是磷酸烯醇式丙酮酸羧激酶。将草酰乙酸脱羧变成磷酸烯醇式丙酮酸,消耗一个高能磷酸键。反应在细胞质和线粒体。
综上,共消耗2分子ATP。
由于丙酮酸羧激酶仅位于线粒体,故细胞质内的丙酮酸必须进入线粒体。而磷酸烯醇式丙酮酸羧激酶细胞质和线粒体内均有,故草酰乙酸可在线粒体中直接转变,也可先转运至细胞质再转变。
草酰乙酸运出线粒体的两种方式:草酰乙酸不能直接透过线粒体内膜。
①经苹果酸转运:线粒体内的苹果酸脱氢酶催化,草酰乙酸——)苹果酸;细胞质内的苹果酸脱氢酶催化,苹果酸——)草酰乙酸。此过程伴随NADH的转运。
②经天冬氨酸转运:线粒体内的谷草转氨酶催化。草酰乙酸——)天冬氨酸;细胞质内的谷草转氨酶,天冬氨酸——)草酰乙酸。此过程无NADH转运。
草酰乙酸由哪种方式转运,主要取决于不同糖异生原料对供氢体的需求。
1,3-二磷酸甘油酸加氢反应需要NADH+H+。
乳酸作为原料,逆反应生成丙酮酸时细胞质就产生了NADH,草酰乙酸经天冬氨酸途径出线粒体。
丙酮酸或生糖氨基酸作为原料,所需的NADH必须由线粒体提供(来自三羧酸循环或脂肪酸β-氧化),草酰乙酸经苹果酸途径出线粒体,以便将NADH运至细胞质利用。

2)果糖-1,6-二磷酸水解为果糖-6-磷酸。
关键酶是果糖二磷酸酶-1。放能反应,不生成ATP。
3)葡糖-6-磷酸水解为葡萄糖。
关键酶是葡糖-6-磷酸酶。
糖异生的关键酶:丙酮酸羧化酶、磷酸烯醇式丙酮酸羧激酶、果糖二磷酸酶-1、葡糖-6-磷酸酶。
4、糖异生的调节
糖异生和糖酵解是方向相反的两条代谢途径,其中3个限速步骤分别由不同的酶催化底物互变,称为底物循环。通常代谢朝酶活性强的方向进行。
Ø 如果催化互变反应的两种酶活性相等,代谢不能向任何方向推进,结果是无谓地消耗ATP而释放热能,形成无效循环。
要促进糖异生就必须抑制糖酵解。二者的反向调节主要是两个底物循环。
1)第一个底物循环调节果糖-6-磷酸与果糖-1,6-二磷酸的互变。
糖酵解,反应耗能;糖异生,反应并无产能。

F-2,6-2P和AMP反向调节第一个底物循环。
F-2,6-2P和AMP既是磷酸果糖激酶-1的别构激活剂,又是果糖二磷酸酶-1的别构抑制剂。这一底物循环的调控最重要。
果糖-2,6-二磷酸是肝内糖酵解和糖异生的主要调节信号。
胰高血糖素减少果糖-2,6-二磷酸,果糖二磷酸酶-1活性增加,糖异生增强而糖酵解减弱,肝内糖原储存增加。
胰岛素作用相反。
2)第二个底物循环调节磷酸烯醇式丙酮酸与丙酮酸的互变。
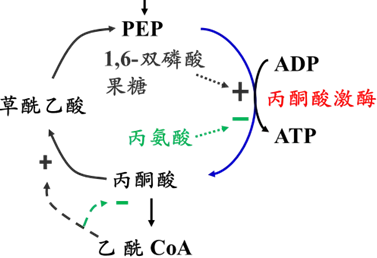
糖酵解,反应产能;糖异生,反应消耗2分子ATP。
u 丙酮酸激酶受别构调节和磷酸化修饰。
别构激活剂:果糖-1,6-二磷酸
别构抑制剂:丙氨酸(饥饿时主要的糖异生原料)
胰高血糖素抑制酶活性(减少F-2,6-2P,进而减少F-1,6-2P;磷酸化丙酮酸激酶抑制其活性)
u 磷酸烯醇式丙酮酸羧激酶受激素诱导的含量调节。
胰高血糖素通过cAMP迅速升高磷酸烯醇式丙酮酸羧激酶的mRNA水平,促进酶蛋白合成,加强糖异生。胰岛素作用相反。
u 乙酰CoA是丙酮酸羧激酶的别构激活剂,也是丙酮酸脱氢酶复合体的别构抑制剂。
生理意义:
1)维持血糖恒定是糖异生最重要的生理作用。
Ø 脑中己糖激酶Km较低,对葡萄糖亲和力较大。
2)糖异生是补充或恢复肝糖原储备的重要途径。
3)肾糖异生增强有利于维持酸碱平衡。预防酸中毒。
5、乳酸循环(Cori循环):
肌收缩(尤氧供不足)通过糖酵解生成乳酸。肌内糖异生活性低,所以乳酸通过细胞膜弥散进入血液后,再入肝,在肝内异生为葡萄糖。葡萄糖释入血液后又可被肌摄取,这就构成了一个循环。
2分子乳酸异生成葡萄糖需消耗6分子ATP。

生理意义:既能回收乳酸中的能量,又可避免乳酸堆积而引起酸中毒。
