
【科普】沼泥鼍迹:浅谈短吻鳄属(Alligator)的化石记录
鼍属Alligator是现存的九属鳄类中的一属,属于鼍科Alligatoridae的鼍亚科Alligatorinae,它还有一个使用更广泛的中文称呼——短吻鳄属(尽管在官方修订的《拉英汉》中“短吻鳄属”是侏儒鳄Osteolaemus的正式中名,但这种叫法显然不怎么被接受。因此本文依然采用“短吻鳄属”作为Alligator的中文名,衍生学名同理,具体请以学名为准)。在一些中文文献中,该属有时也被称为猛鳄或钝吻鳄。
现存的短吻鳄属成员只有两种,包括美国东南部的密西西比短吻鳄Alligator misisipiensis和我国长江流域的扬子鳄Alligator sinensis:


这两种短吻鳄的地理位置分布差异相当大。因此,作为不具备长途跨海迁徙能力的淡水动物,短吻鳄在其历史上的分布显然要较现代广得多。同时短吻鳄属也是鳄目这九个现存属中最古老的一个,其可靠的化石记录可以追溯至3720-3390万年前的晚始新世普利亚本期,化石产出相当丰富,且已经发现并确认了五至七个化石物种。《中国古脊椎动物志》第二卷第四册中提供的短吻鳄科(书中称为钝吻鳄科)的鉴别特征为:
吻部宽而扁平,两侧缘平行,具雕饰纹。外鼻孔被前颌骨和鼻骨所围,鼻骨形成鼻孔内纵向中隔。泪骨小于前额骨,鼻骨的后端不与泪骨相接,它们被相连的前额骨、上颌骨所隔开。眼眶大,其前缘不超过上颌第十一齿。顶骨直达颅骨后缘,上枕骨不参与或仅少许构成颅骨的顶面。头骨腹面左、右前颌骨紧密接合,每侧5齿。腭骨前伸最远达第七上颌骨齿。


笔者在撰写本文时搜集了一些相关的研究文献,在此简单的介绍一下短吻鳄属的化石种和现存种的化石记录。

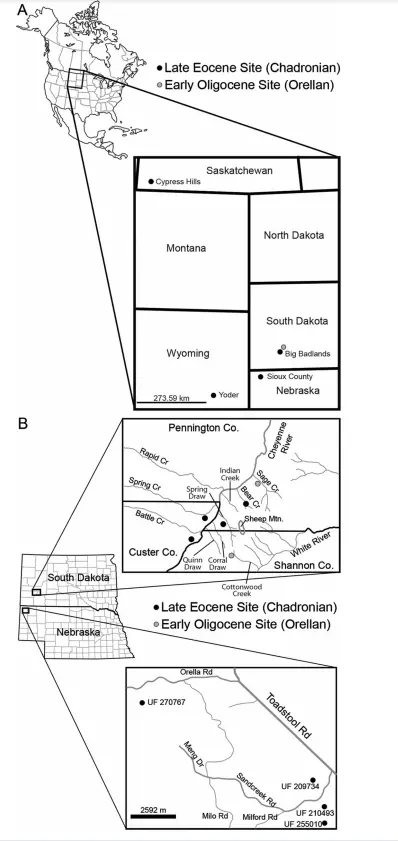
北美大陆是短吻鳄科的发祥地,也是短吻鳄属的故乡。已知最早出现的短吻鳄属成员是前鼻短吻鳄Alligator prenasalis,一种小型的、较为基干的史前短吻鳄,目前已经发现了许多化石材料和疑似化石,出土于晚始新世到早渐新世的美国北部。




前鼻短吻鳄的化石分布在著名的白河恶地(White River Badlands),主要是南达科他州一带的查德伦组Chadron Formation和布鲁尔组Brule Formatio地层。伊万·怀廷(Evan Whiting)和亚历山大·哈斯汀(Alexander Hasting)等人2015年的研究报道了一些出土于晚始新世内布拉斯加州西北部的短吻鳄属头骨和下颌材料,地层同样是查德伦组,也可能是前鼻短吻鳄,只是缺乏有效的诊断特征。另外,一些来自怀俄明州和加拿大萨斯喀彻温省的短吻鳄材料也被认为可能属于该种。


对前鼻短吻鳄这一物种的发现和研究始于一百多年前。它在1904年最初命名时被作为鳄属Crocodylus的一员,即前鼻“鳄”Crocodylus prenasalis,1926年才划入短吻鳄属。还有同地区1916年命名的“维氏似凯门鳄”Caimanoidea visheri和1931年命名的“里氏异颌鳄”Allognathosuchus riggsi,这两个物种最终都被分入短吻鳄属,合并入前鼻凯门鳄种。


前鼻短吻鳄有着较宽的鼻部,外观与现代的密河短吻鳄相差不大。但其体型要小得多,基本与扬子鳄相仿。其头骨尺寸可达243.5mm,对应体重据算约为17.4kg。




白河恶地发现了种类丰富的哺乳类,其中包括渐新马Mesohippus、先兽Poebrotherium等有蹄类食草动物,也不乏伪剑齿虎Hoplophoneus、恐齿猫Dinictis和鬣齿兽Hyaenodon等活跃的食肉者,不过限于笔者对古兽的几乎一无所知,就暂且不详细记述。另外,当地出土的各类蜥蜴鳞龙也相当多,还有几种鱼类、龟类和蟾。前鼻短吻鳄很可能经历了始新世末当地环境由(查德伦组)炎热、湿润转向(布鲁尔组)干燥、较冷的过程,并受其影响逐渐缩减灭绝。
另一种较早期和原始的短吻鳄属成员是麦氏短吻鳄Alligator mcgrewi,该种1941年被卡尔·施密特(Karl Schmidt)描述命名,种加词是为了纪念标本的收集者保罗·麦格雷夫(Paul McGrew)。麦氏短吻鳄最初只确认了两个标本,即正模F.M. No. P26242(FMNH P26242)和参考标本F.M. No. P26281。正模是一个几乎完整的颅骨,并带有接近完整的下颌支。而参考标本的保存部位则包括两块上颌骨碎片,一些相关牙齿和其他小骨片。后来又发现了许多完整或不完整的头骨和下颌材料,这些材料在布莱恩·马龙(Brian Malone)1979年的论文里有所提及。


麦氏短吻鳄模式产地位于内布拉斯加州西北部的阿莱托默里克斯矿场(Aletomeryx Quarry),地层为海明福群Hemingford Group的流水组Runningwater Formation(又叫马尔斯兰组Marsland Formation,施密特将其写作"Marseland Formation",疑系拼写错误),年代为早中新世(大部分短吻鳄种类都出现在中新世)。麦氏短吻鳄所在的流水组也发现了各种各样的史前兽类,包括好几种犬类以及鼠、鼹鼠、鼠兔之类的啮齿动物。另外,周边的其他地层也有该物种出土,如绵羊溪组Sheep Creek Formation和蛇溪组Snake Creek Formation。


麦氏短吻鳄被认为是“后来所有短吻鳄的祖先,包括现存的两个种”。在大多数的系统发生学研究中,麦氏短吻鳄和前鼻短吻鳄常被作为姊妹群,构成了一个原始的短吻鳄属分支,被排除在短吻鳄属冠群之外。目前发现的麦氏短吻鳄体型很小,F.M. No. P26242的颅骨长度只有145.2mm(1942年命名论文),DCL为139mm(2019年论文补充材料提供),比一般的扬子鳄还要更小。麦氏短吻鳄的头骨宽度大于其长度,可能是吻部最钝的短吻鳄,吻形有些接近扬子鳄,可能倾向以硬壳软体动物为主食。

同样在阿莱托默里克斯矿场还发现了一些短吻鳄相似种cf. Alligator化石,伊万·怀延和杰森·海德(Jason Head)在2020年的论文中就对这些材料进行了描述。涉及的标本有UNSM 135037和UNSM 135036两个,包括很多零散的颅骨和躯干材料。因为缺乏与现有种类区分开的自有特征,这些材料很可能也属于麦氏短吻鳄。


奥氏短吻鳄Alligator olseni命名于1942年,命名者是西奥多·怀特(Theodore White),种加词纪念罗塞尔·奥尔森(Russel Olsen)。它的化石已发现于佛罗里达州和德克萨斯州的几处,年代也是中新世早期。模式产地是佛罗里达阿拉楚阿组Alachua Formation(或霍桑组Hawthorn formation,这两个地层名称的使用存在争议)的托马斯农场(Thomas Farm),该地出土了两个标本:正模标本MCZ1887和副模标本MCZ1888,分别是一个保存完好的颅骨和来自一个更大个体的左侧下颌。



后来,一些更完整的标本也被从该地点挖掘出来。古生物学家和志愿者在托马斯农场的采集工作目前依然在定期进行。克里斯多夫·布罗楚(Christopher Brochu)和丹尼尔·斯奈德(Daniel Snyder)等学者曾报道过一些新发现的奥氏短吻鳄标本,包括更丰富的头后材料。


奥氏短吻鳄方骨的宽度仅仅略长于全长的一半,下颌齿列的长度约等于整体长度的一半。头骨轮廓和尺寸比例接近于密河鳄。奥氏短吻鳄被描述为“一个成熟期的小型物种”,并可能是更衍化的短吻鳄的祖先,是介于短吻鳄属原始成员和衍化成员之间的中间物种。根据2011年的报道,田纳西州东北部中新世晚期至上新世早期地层发现了一种尚未被描述的小型短吻鳄,初步分析表明,这种短吻鳄具有奥氏短吻鳄和一些更衍化短吻鳄的特征。
1957年,沃尔特·奥芬博格(Walter Auffenberg)报道了在佛罗里达州的香榧组Torreya Formation地层出土的一些奥氏短吻鳄材料,涉及的馆藏编号包括FGS V-3785, 1440, 1439, 1437, 2777, 2768,1436,5315和4168,保存部位包括许多颅骨、下颌和头后骨架等残片:

1994年,巴里·奥尔布赖特(Barry Albright)又记录了来自德克萨斯州弗莱明组(Fleming Formation)的奥氏短吻鳄,包括牙齿和其他零碎的材料。不过德克萨斯的奥氏短吻鳄材料比佛罗里达的更加古老,而且体型更大,可能属于一个新物种。在这些材料被回顾之前,它们是否属于奥氏种是值得怀疑的。然而,如果这些材料确实属于奥氏短吻鳄,那么奥尔布赖特的记录将为该物种的体型及地理地史分布提供显著的范围扩展。


“汤氏”短吻鳄Alligator 'thomsoni'来自内布拉斯加州中新世中期奥尔柯特组Olcott Formation地层的特洛伊采石场(Trojan Quarry)。目前只报道了一例确认的化石记录,即1921-1922年间阿尔伯特·汤姆森(Albert Thomson)采集的头骨和下颌,这个物种最终也以他的姓氏命名。1923年,查尔斯·穆克(Charles Mook)描述并命名了这一新种(此人也是最早研究我国鳄类化石的学者)。这一物种的有效性当前还存在争议。
汤姆森采集到的标本包括正模AMNH 1736——一个“保存完好的、中等大小的颅骨”以及三个副模:AMNH1727、1738和1739,它们的保存部位都是下颌支。AMNH的尺寸接近现代密河短吻鳄,DCL为320mm,颅长可达36厘米,比此前的短吻鳄种类要大得多。





“汤氏”‘短吻鳄的口鼻部通常比密河短吻鳄更短,但没有明显的缝合差异。在布莱恩·马龙1979年的论文中被划为现代密西西比短吻鳄的初级同物异名,同时也将密河短吻鳄的历史前推到了1400万年前。克里斯多夫·布罗楚依照关节动脉舌孔的位置和口腔颌间前孔的存在,对这一分属提出怀疑(不过他在1999年的论文中似乎将学名误拼成了"Alligator thompsoni")。他还提到在德克萨斯也发现了“汤氏”短吻鳄相似种cf. Alligator thomsoni的颅骨材料,馆藏编号为TMM 3 1219-233。

同样在1979年被布莱恩·马龙划为密河短吻鳄异名的还有穆克1946年命名的梅氏短吻鳄Alligator mefferdi 。梅氏短吻鳄出土于上新世内布拉斯加州的灰穴组Ash Hollow Formation,正模是1941年夏天拉尔夫·梅弗尔德(Ralph Mefferd)采集的AMNH 7016,包括“保存完好的颅骨和下颌,一个脊椎的神经棘,一些鳞片和一块舌骨”。按照惯例,新物种也以发现者的姓氏命名。

穆克最初认为梅氏短吻鳄是扬子鳄的近亲,但其实它在形态上反而和密河短吻鳄高度接近。因此同汤氏种一道被马龙作为后者的异名。布罗楚在1999年的论文中再次肯定了梅氏种和密河种“极端地(extremely)”相似,但指出两者可以通过perforate splenial(不知道怎么翻译)区分开。梅氏种的splenial被排除在下颌联合之外,但仍然保留了一个anterior foramen intermandibularis oralis,这一特征从未在密河短吻鳄上出现过。因此,梅氏短吻鳄重新被视为一个有效物种,而且是密河短吻鳄的已知近亲,它们间的关系应当比现存的两个短吻鳄属物种更近。因此,梅氏种被认为是“冠群(crown)”的短吻鳄属成员。
它可能也是最早的冠群短吻鳄——斯奈德在2007年的论文中报道了来自在佛罗里达州阿拉楚阿组Alachua Formation(或霍桑组Hawthorn formation)的托马斯农场(Thomas Farm)的梅氏短吻鳄相似种Alligator cf. mefferdi材料——这里也就是前面提到过的奥氏短吻鳄的模式产地,年代为中新世早期。设计的材料包括一个下颌残片和很多头后材料。如果这个记录真的属于梅氏短吻鳄,那么它的地史分布将大大拓宽,同时可能和体型娇小的原始亲戚奥氏短吻鳄共享栖息地。不过在伊万·怀廷等人2016年的报道里,这些材料被重新分析并认定为密河短吻鳄相似种Alligator cf. mississippiensis
海尔短吻鳄Alligator hailensis是杰里米·斯托特(Jeremy Stout)在2020年的论文新近描述命名的物种。海尔短吻鳄得名于其发现地,佛罗里达州中北部的海尔镇(Haile)的7C和7G两个采集点。正模标本UF 224688,保存部位包括完好的颅骨、下颌和很多零碎的骨甲、躯干、四肢等头后材料。两个采集点共出土了数十上百个被划分为海尔短吻鳄的标本,大都比较零碎:



海尔短吻鳄被描述为一种“大型的”短吻鳄属成员,其体型堪与现存的密河鳄相当。海尔短吻鳄相当衍化,其年代和骨骼形态介于地质学上更古老的梅氏短吻鳄和现代密西西比短吻鳄之间,因此是一种过渡化石。
另外,海尔短吻鳄还出土了一些罕见的粪化石:

东亚也有数量相当的短吻鳄属化石记录。除了存活至今的扬子鳄之外,鲁短吻鳄Alligator luicus也是一种产自我国的短吻鳄属成员。目前该种只记录了一个标本,即正模SWPM(LPM 850001),一个接近完整的头骨、破碎的下颌和部分头后骨架。该标本现存于中国古动物馆,笔者也有幸得以近距离观察。鲁短吻鼍命名于1987年,命名者是中国科学院古脊椎动物与古人类研究所的李锦玲和山东临朐古生物博物馆的王宝忠。



鲁短吻鳄产于山东临朐县尧山山旺盆地,地层为山旺组Shanwang Formation,年代为中中新世,属于著名的山旺生物群(Shanwang Biota)。山旺生物群已经出土了种类繁多、颇具代表性的鱼类和哺乳类化石,包括近无角犀Plesiaceratherium、原古鹿Palaeomeryx、犬熊Amphicyon等等。
鲁短吻鳄有着短小的头骨,“表面颅刻纹发育。吻部长度小干宽度,也小于头骨其余部分的长度。上颌骨与前额骨、鼻骨与泪骨互成对角接触。眶前嵴不发育;上颞凹较宽大,呈长椭圆形。”,LPM 850001的颅骨长度据测为105mm,体长约60cm。如果以扬子鳄的生长为标准,该标本的体型似乎代表着一个尚未达到性成熟的较大的幼年个体。其他一些解剖学特征也支持鲁短吻鳄正模尚未成年。对该种的研究主要集中在头部,头后的躯干和四肢材料虽然完整,但保存状态不佳,多数地方难以辨识。

鲁短吻鳄未被纳入系统发生学研究,其与扬子鳄和其他短吻鳄属成员的关系尚不明确。不过鲁短吻鳄和扬子鳄的形态存在确凿的差异,而整体上似乎更具原始短吻鳄属的特征(原文认为它和麦氏短吻鳄最为相似)。因此,鲁短吻鳄的祖先可能并非与扬子鳄一道迁徙到东亚。或许,短吻鳄属曾不止一次向大洋彼岸扩散。一般认为短吻鳄是通过白令路桥到达亚洲的,但具体的时间仍然有待确认。
现存的密西西比短吻鳄和扬子鳄各自都有很多化石记录,但其中大都是一些意思材料。此前的分子生物学研究指出这两者的分化时间在5800-3100万年前。
密河种最早的相关化石记录可能是古新世南卡罗莱纳黑明戈群(Black Mingo Group)出土的一个皮内成骨,1998年被描述并划分为“密河短吻鳄相似种”Alligator cf. mississippiensis。不过考虑到材料很少、只有文字描述,而且年代实在过早,这一归属十分可疑,基本可以确定是不可靠的。

伊万·怀廷等人在2019年也报道了出土于佛罗里达早中新世密河短吻鳄相似种Alligator cf. mississippiensis,就是在2007年被称为梅氏短吻鳄相似种的那些材料,在上文中已经记述过。如果这些材料确切属于密河短吻鳄,那么这将是该物种年代最早的材料之一(另外还有之前的“汤氏”短吻鳄)




斯蒂芬·梅(Steven May)等人在2019年报道了德克萨斯州南部拉帕拉河动物群(Lapara Creek Fauna)的密河短吻鳄相似种材料,包括大量牙齿和骨甲,以及一些头部和头后骨骼:






尽管现代扬子鳄的出没区域仅限于长江下游狭小的栖息地间,但丰富的化石材料使人不得不相信它们曾有大得多的分布范围,甚至不只限于中国。扬子鳄最古老的化石记录可能是来自中中新世泰国呵叻府的一个鳄类头骨DMR-BSL2011-2,该标本2011年被朱利安·克劳德(Julien Claude)等人报道,并划为扬子鳄相似种Alligator cf. sinensis。其外形短而粗壮,长度可达25cm——这可能对应了一个2-3米的相当大的个体。


而在已确认的扬子鳄化石材料中,最古老的则出土自日本晚上新世。包括左方骨LBM 0142000027,左齿骨 LBM 0142000753,左隅骨LBM 0142000756和左关节骨LBM 0142000756(这些左下颌材料属于同个个体)。这些扬子鳄体型较大,尺寸和现代种群相当。

东亚的板块构造和地理历史表明,短吻鳄可能在2500万年前之前或1000万年前之后扩散到日本,但由于日本列岛的半孤立条件和上更新世的气候恶化,最终在日本区域性灭绝。
我国国内的扬子鳄(包括扬子鳄相似种)化石分布相当零散,遍及山西、山东、安徽、河南、浙江、广东和台湾,其中主要是牙齿和骨甲。除了广东、安徽记录的两例来自中更新世之外,其他的均来自新石器时代。
唯一保存完好的头骨记录产自台湾,出土位置在台湾和澎湖岛间澎湖海峡的澎湖隧道中,编号为NMNS006394-F051722。




本文到此为止。由于涉及信息量巨大,笔者能力有限,难免疏漏,还望补充指正。
【参考文献】
{1} Rio&Mannion 2021:Phylogenetic analysis of a new morphological dataset elucidates the evolutionary history of Crocodylia and resolves the long-standing gharial problem
{2}Snyder 2007:Morphology and systematics of two Miocene Alligators from Florida, with a discussion of Alligator Biogeography
{3}Stout 2020.:New early Pleistocene Alligator (Eusuchia: Crocodylia) from Florida bridges a
gap in Alligator evolution
{4}Whiting and Hasting 2015:First Fossil Alligator from the Late Eocene of Nebraska and the Late Paleogene Record of Alligators in the Great Plains
{5}Stout 2021:Some thoughts on Interspecific Mandibular morphology in fossil and modern Alligator
{6}Schmidt 1941:A new fossil alligator from Nebraska
{7}Loomis 1904:Two new river reptiles from the Titanothere beds
{8}Mook 1923.:A new species of alligator from the Snake Creek Beds
{9}Brachu 1999:Phylogenetics, Taxonomy, and Historical Biogeography of Alligatoroidea
{10}Malone 1979:The Systematics, Phylogeny and Paleobiology of the Genus Alligator
{11}Whiting&Head 2020:Neogene crocodylians from the Central Great Plains: spatiotemporal histories and relationships with regional climatic and environmental changes
{12}White 1942:A New Alligator from the Miocene of Florida
{13}Mook 1946:A new Pliocene alligator from Nebraska
{14}Li&Wang 1987:记山东山旺钝吻鳄(Alligator)一新种
{15}May 2019:The Lapara Creek Fauna: Early Clarendonian of south Texas, USA
{16}Erickson 1998:Crocodilians of the Black Mingo Group (Paleocene) of the South Carolina Coastal Plain
{17}Whiting et al.2016:Cranial Polymorphism and Systematics of Miocene and
Living Alligator in North America
{18}Claude et al.2011:Neogene reptiles of northeastern Thailand and their paleogeographical signifificance
{19}Iijima et al. 2016:The oldest record of Alligator sinensis from the Late Pliocene of Western Japan, and its biogeographic implication
{20}Shan et al.2013:The first fossil skull of Alligator sinensis from the Pleistocene, Taiwan, with a paleogeographic implication of the species
